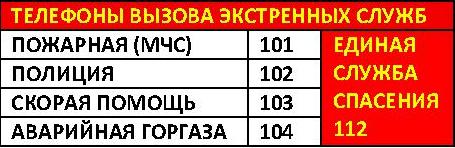


2011-2017 © МБУЗ ГКП № 7, г.Челябинск.
Ознакомиться с примером...
Имеются противопоказания. Необходимо проконсультироваться со специалистом.«Фосфоглив» – пример современного комбинированного препарата, способного помочь на всех стадиях поражения печени:
Какие средства в этом помогают?
Ознакомиться с примером...
Печень — крайне сложный орган, отвечающий, помимо всего прочего, за фильтрацию крови и расщепление вредных веществ — как поступающих в организм извне, так и являющихся побочными продуктами обмена веществ. Она нуждается в бережном и внимательном отношении. Часто приходится слышать, что положительный эффект даже на фоне здорового образа жизни может оказать периодическое очищение печени.
Так называемая чистка от шлаков овеяна множеством мифов. Чтобы не навредить своему организму и не перегрузить печень биологически активными и далеко не всегда безвредными веществами, необходимо подходить к вопросу комплексно, не увлекаясь поиском какого-то одного быстродействующего средства. Давайте вместе посмотрим, какие существуют пути решения проблемы.
Шлаки — это конечные продукты обмена веществ. Токсины — вредные компоненты, попадающие в организм извне или образующиеся в нем. И те и другие, накапливаясь в нашем теле, вызывают внутреннюю интоксикацию, которая может проявляться в повышенной утомляемости, головных и суставных болях, кожных высыпаниях, аллергии, повышении температуры тела. Кроме того, вредные вещества образующиеся в печени (жиры) или попадающие в нее извне (яды, алкоголь, некоторые лекарства) оказывают повреждающее действие и на саму печень, вызывая ее ожирение, воспаление, фиброз и, как следствие, нарушений функций. В результате печень плохо справляется со своими задачами по своевременному выводу ядовитых веществ, и все эти негативные процессы усиливаются.
Стоит сказать, что проблемы с функционированием печени в той или иной степени присущи практически каждому взрослому жителю мегаполиса в силу техногенных, экологических, социальных факторов, которые усугубляются вредными привычками — приемом алкоголя, курением, употреблением фастфуда.
Нередко предлагаются различные методики «чистки» печени от шлаков и токсинов. Сразу стоит сказать, что «чистки» печени не существует в принципе, а все предлагаемые методики в лучшем случае являются неизученными, а в худшем – шарлатанством.
В Интернете вы найдете немало нумерованных списков с «методами» и «способами» очищения печени. Скажем прямо, сама постановка этого вопроса — абсурд. Печень не губка, которую можно выжать, не тряпка, которую можно постирать, и не фильтр в нашем бытовом понимании. Печень сама отвечает за собственную самоочистку, равно как и за очищение всего организма в целом. Если она справляется с возложенной на нее природой задачей, лучшей помощью будет соблюдение правил здорового образа жизни. При нарушении функций по тем или иным причинам необходимы коррекция образа жизни, питания и, не редко, лекарственная терапия. Если же печень перестает выполнять свою основную функцию требуется серьезное медикаментозное лечение. Чтобы узнать о состоянии печени необходимо проконсультироваться с терапевтом или гастроэнтерологом (гепатологом), пройти диагностические процедуры (УЗИ, биохимический и серологический анализы крови).
Кроме неэффективных методов «чистки» печени , не редко предлагаются столь же сомнительные процедуры подготовки к ней.
Так рекомендуют несколько дней соблюдать вегетарианскую диету, очистить кишечник с помощью клизмы или ежедневного приема в течение нескольких недель меда, раствориенного его в теплой воде.
Ниже описано несколько народных средств», применив которые можно получить «чистую» работоспособную печень,. Не стоит поддаваться соблазну и применять не глядя какой бы то ни было рецепт из Интернета типа «очистка печени». Отнеситесь к проблеме по-взрослому.
Народные средства
Народная медицина предлагает массу средств для очищения печени. Например, мед расторопши, тыквенные семечки и масло, орехи и другие. Кроме недоказанной эффективности они могут иметь и противопоказания. Так, мед противопоказан при сахарном диабете.
В отдельных случаях, некоторыми врачами упоминается такой метод, как тюбаж — прием желчегонных средств (минеральной воды, травяных отваров, льняного или оливкового масла с лимонным соком) с одновременным прогреванием области печени обыкновенной грелкой с частотой 1–2 раза в неделю в течение месяца. Смысл тюбажа — вызвать сокращение желчного пузыря, чтобы предотвратить застой желчи. Агрессивное проведение процедуры может привести к рвотному рефлексу. В целом отношение к данной процедуре официальной медицины скорее отрицательное. Более того, она относится не столько к печени, сколько желчевыводящим путям. Можно услышать от лжеврачей такие советы, как, например, питье свежего оливкового масла первого отжима со свежевыжатым лимонным соком частями в течение дня вплоть до возникновения тошноты. Первая половина предложения звучит жизнеутверждающе, но вторая… — при ее реализации вы можете получить повреждение (вплоть до разрыва) желчного пузыря, особенно, если в нем находятся камни. И наконец, на любой жир, а не только на «экстра-вёрджин» оливковое масло, желчный пузырь реагирует сокращением — чем больше дозировка, тем сильнее спазм — но если цель заключается в рвотном рефлексе, то не имеет никакого значения, чем он вызван. И еще: в домашних условиях стимулировать желчный пузырь следует естественным образом и только перед едой или во время нее — желчь должна реализовать свою функцию, а не дополнительно отравлять организм. И никогда не прибегайте к прогреванию печени без строгих и однозначных указаний врача: вы можете спровоцировать внутреннее кровотечение, которое, в свою очередь, приведет к летальному исходу. Категорически запрещено прогревание области печени при наличии гастрита, язвы и воспалительных процессов поджелудочной железы.
Диета для восстановления печени
Стоит морально подготовиться к тому, что восстановление печени потребует от вас серьезных волевых усилий. Хорошая новость состоит в том, что диета ни в коем случае не должна быть «голодной», вы будете страдать скорее от отсутствия «вкусненького».
Итак, на период хотя бы одной недели необходимо исключить из рациона белый хлеб, сахар, острое, жареное, жирное, соленое. Категорически противопоказан алкоголь и табакокурение. Рекомендованы сухари, овощные супы, нежирное мясо и птица, злаковые, некислые овощи и фрукты, мед, чай, отвар шиповника, свежевыжатые соки (особенно овощные). Например, положительное влияние на печень оказывает коктейль из соков свеклы, яблока, огурца, лимона, приправленный чайной ложкой оливкового масла. В день достаточно выпивать 1/3 стакана этой смеси натощак. Лучше всего питаться по утвержденной схеме, пять раз в день и небольшими порциями. Так называемые диеты № 5 (№ 5а), № 7, № 15 и их комбинации назначают пациентам при проблемах с печенью уже не одно десятилетие, они помогают и при очищении печени. Особенностями медицинских диет являются дробное питание (6–7 раз в день), приготовление паровой и протертой пищи, высокое содержание белка. Пациентам с хроническим гепатитом стол № 5 прописывают на постоянной основе. На этот же период имеет смысл воспользоваться бессолевой диетой № 7.
В питании вы должны соблюдать несколько неукоснительных правил:
Роль правильного питания в восстановлении печени сложно переоценить. Конечно, психологически тяжело отказаться от любимых блюд на длительный срок. Однако освобождение от тяжести в животе, снижение утомляемости, посвежевший внешний вид через несколько дней соблюдения диеты и приема препаратов помогают пациентам выдержать это испытание. Если вы мысленно отстранитесь от надуманной необходимости во вредной пище и от желания себя побаловать, вскоре придет осознание, что вы можете обходиться без нее сколь угодно долго.
Лекарственные препараты
Для восстановления нормальной работы печени назначаются преимущественно два типа лекарственных средств — гепатопротекторы и желчегонные препараты, обеспечивающие защиту и восстановление клеток печени, а также нормальный отток желчи. В гепатопротекторах используются такие действующие вещества, как эссенциальные фосфолипиды, флавоноиды, глицирризиновая кислота и другие. Желчегонные препараты создаются на основе медицинской желчи животного происхождения и/или растительных компонентов, усиливающих секрецию желчного пузыря. Следует понимать, что все органы ЖКТ находятся в тесной взаимосвязи, а значит — сбой в работе печени может вызываться нарушениями в функционировании желудка, кишечника, различных желез кроме печени. Поэтому лекарственная терапия строится на принципах комплексности.
Константа стабильности, образования или связывания - это тип константы равновесия, используемой для образования металла комплексы в растворе. Точно, константа стабильности применима для измерения силы взаимодействия между лигандами и ионами металлов, которые участвуют в комплексообразовании в растворе [11].Как правило, эти 1-4 уравнения выражаются следующим образом:
Металл + Лиганд-Металл-ЛигандK1 = MLMLE1
Металл + ЛигандыandМеталл + Лиганд2K2 = ML2MLLE2
Металл + Лиганд3⇆Металл + Лиганд3K3 =
ML3MLМеталл + Ligandn − 1 + L⇆Металл + Лиганд − nKn = MLnMLn − 1LE4
K 1 , K 2 , K 3 ,… K n являются константами равновесия, и их также называют ступенчатые константы устойчивости. Образование комплекса металл-лиганд -n также можно выразить как константы равновесия с помощью следующих шагов:
Металл + лиганд → B1Металл-лиганд, β = MLMLE5
Металл + 2Лиганд → B2Metal-лиганд2, β2 = ML2ML2E6
Таким образом, Metal + nLigand → BnMetal-лигандLn, βn = MLnMLnE7
β 1 , β 2 , β 3 ,… β n являются константами равновесия, и эти константы равновесия известны как общая стабильность константы или общее образование.β n называется n-й совокупной или общей константой пласта [12]. Любые комплексы металлов будут иметь большую стабильность, если их константа устойчивости будет иметь более высокое значение. Иногда значения 1 / k являются альтернативными значениями константы устойчивости, и теперь это называется константой нестабильности. Log 10 K 1 , log 10 K 2 … log 10 K n и log 10 β n являются способами, которые выражают ступенчатую и кумулятивную константы устойчивости.
Параметры K и β связаны между собой, и они выражаются в следующий пример:
β3 = ML3ML3E8
Теперь числитель и знаменатель умножаются вместе с использованием [металл-лиганд] [металл-лиганд 2 ], и после перегруппировки мы получаем следующее уравнение:
β3 = ML3ML3 × MLML2MLML2 = ML1ML × ML2MLL × ML3ML2L = K1xK2xK3E9
Таким образом, βn = ML1ML × ML2MLL….MLnMLn − 1L = K1xK2… KnE10
Теперь мы выразили это следующим образом:
βn = ∑n = 1n = nKnE11
Из приведенного выше соотношения ясно, что общая константа устойчивости β n равна произведение последовательных (то есть ступенчатых) констант устойчивости, K 1 , K 2 , K 3 ,… K n . Другими словами, это означает, что значение констант устойчивости для данного комплекса фактически состоит из ряда ступенчатых констант устойчивости.Термин «стабильность» используется без уточнения для обозначения того, что комплекс существует в подходящих условиях и что его можно хранить в течение значительного периода времени. Термин стабильность обычно используется, потому что координационные соединения стабильны в одном реагенте, но диссоциируют или растворяются в присутствии другого реагента. Также возможно, что термин стабильность может упоминаться как действие тепла или света или соединения. Устойчивость комплекса [13] выражена качественно в терминах термодинамической и кинетической устойчивости.
В химической реакции химическое равновесие - это состояние, в котором концентрация реагентов и продуктов со временем не изменяется. Часто это условие возникает, когда скорость прямой реакции становится такой же, как скорость обратной реакции. Стоит отметить, что скорости прямой и обратной реакции на этой стадии не равны нулю, а равны.
Если водород и йод хранятся вместе в молекулярных пропорциях в закрытом технологическом сосуде при высокой температуре (500 ° C), начинается следующее действие:
h3 + I2 → 2HIE12
В этой операции йодид водорода образуется путем объединения водорода и йода, а количество йодистого водорода увеличивается со временем.В противоположность этому действию, если чистый газообразный йодид водорода нагревают до 500 ° С в реакции, соединение растворяется обратным действием, что вызывает растворение йодида водорода в водороде и йоде, и соотношение этих продуктов со временем увеличивается , Это выражается в следующей реакции:
2HI → h3 + I2E13
Для образования хелатов металлов термодинамический метод дает очень важную информацию. Термодинамика - очень полезный метод, позволяющий различать энтальпические и энтропийные эффекты.Сила связи полностью зависит от энтальпического эффекта, и это не имеет никакого значения во всем решении в порядке / беспорядке. Основываясь на термодинамике, хелатный эффект ниже может быть лучше всего объяснен. Изменение стандартной свободной энергии Гиббса для константы равновесия - это отклик:
ΔG = −2,303RTlog10β.E14
Где:
R = газовая постоянная
T = абсолютная температура
При 25 ° C,
ΔG = ( - 5,708 кДж, моль -1 ) · log β.
Термин энтальпии создает свободную энергию, т.е.
ΔG = ΔH – TΔSE15
Для комплексов металлов термодинамическая стабильность и кинетическая стабильность являются двумя интерпретациями константы устойчивости в растворе. Если реакция переходит от реагентов к продуктам, это относится к изменению ее энергии, как показано в приведенном выше уравнении. Но за реактивность, кинетическая стабильность отвечает за эту систему, и это относится к видам лигандов [14].
Стабильный и нестабильный - термодинамические термины, а лабильный и инертный - кинетические.Как правило, те комплексы, которые полностью реагируют в течение примерно 1 минуты при 25 ° С, считаются лабильными, а те комплексы, для реакции которых требуется больше времени, считаются инертными. [Ni (CN) 4 ] 2- является термодинамически стабильным, но кинетически инертным, поскольку он быстро обменивается лигандами.
Комплексы металлов [Co (Nh4) 6 ] 3+ и другие типы таких комплексов кинетически инертны, но они термодинамически нестабильны. Мы можем ожидать, что комплекс разлагается в присутствии кислоты немедленно, потому что комплекс термодинамически нестабилен.Скорость разложения в кислотном растворе составляет порядка 10 25 . Следовательно, оно термодинамически нестабильно. Однако с комплексом ничего не происходит, если его хранить в кислом растворе в течение нескольких дней. При рассмотрении устойчивости комплекса всегда должно быть указано условие. При каких условиях должен быть указан комплекс, который является стабильным или нестабильным, например, кислотный, а также основное состояние, температура, реагент и т. Д.
Комплекс может быть стабильным по отношению к определенному состоянию, но по отношению к другому.Вкратце, стабильный комплекс не должен быть инертным и аналогичным образом, а нестабильный комплекс не должен быть лабильным. Это мера степени образования или превращения комплекса при заданном наборе условий в равновесии [15].
Термодинамическая стабильность играет важную роль в определении прочности связи между металлическими лигандами. Некоторые комплексы стабильны, но как только они вводятся в водный раствор, видно, что эти комплексы влияют на стабильность и распадаются.Для примера возьмем комплекс [Co (SCN) 4 ] 2+ . Ионная связь этого комплекса очень слабая и быстро разрушается, образуя другие соединения. Но когда [Fe (CN) 6 ] 3- растворяется в воде, он не тестирует Fe 3+ с помощью какого-либо чувствительного реагента, что показывает, что этот комплекс более стабилен в водном растворе. Таким образом, указывается, что термодинамическая стабильность связана с энергией связи металл-лиганд, константой стабильности и другими термодинамическими параметрами.
Этот пример также предполагает, что термодинамическая стабильность относится к стабильности и нестабильности комплексов. Измерение степени превращения одного вида видов в другой вид может быть определено по термодинамической стабильности до достижения равновесия. Например, тетрацианоникелат представляет собой термодинамически стабильный и кинетически лабильный комплекс. Но пример катиона гексаамина кобальта (III) прямо противоположен:
CoNh463 ++ 6h4O + → Coh3O63 ++ 6Nh5 + E16
Термодинамика используется для выражения разницы между стабильностью и инерцией.Для стабильного комплекса большие положительные свободные энергии были получены из реакции ΔG 0 . ΔH 0 , стандартное изменение энтальпии для этой реакции, связано с константой равновесия β n уравнением термодинамики скважины:
ΔG0 = -RTlnβE17
ΔG0 = ΔH0-TΔS0E18
Для подобных комплексов Значения ΔS 0 для различных ионов одного и того же заряда конкретного переходного ряда и конкретного лиганда не будут существенно отличаться, и, следовательно, изменение значения ΔH 0 будет связано с изменением значений β n .Таким образом, порядок значений ΔH 0 также является порядком значения β n .
Кинетическая стабильность относится к скорости реакции между ионами металла и лигандом, протекающей в равновесии или используемой для образования комплексов металлов. Чтобы принять решение о кинетической устойчивости любых комплексов, время является фактором, который играет важную роль для этого. Он имеет дело между скоростью реакции и механизмом реакции этого комплекса металла.
Как мы обсуждали выше в термодинамической стабильности, кинетическая стабильность относится к комплексам, в которых комплекс является инертным или лабильным. Термин «инертный» использовался Tube для термически стабильного комплекса, а для реактивных комплексов использовался термин «лабильный» [16]. Природный хлорофилл является примером полидентатного лиганда. Этот комплекс является чрезвычайно инертным благодаря обмену иона Mg 2+ в водных средах.
Характер центрального атома комплексов металлов, размерность, степень его окисления, электронное строение этих комплексов и многие другие свойства комплексов зависят от константы устойчивости.Некоторые из следующих факторов описаны ниже.
В координационной химии комплексы металлов образуются в результате взаимодействия ионов металлов и лигандов. Для соединений этого типа ионы металлов являются координационным центром, а лиганд или комплексообразующие агенты ориентированы вокруг него. Эти ионы металлов в основном являются переходными элементами. Для определения константы стабильности некоторые важные характеристики этих комплексов металлов могут быть такими, как приведенные ниже.
Лиганды ориентированы вокруг центральных ионов металлов в комплексах металлов. Размеры этих ионов металлов определяют количество видов лигандов, которые будут присоединяться или ординатироваться (дательный ковалент) в образовании связи. Если размеры этих ионов металлов увеличиваются, стабильность координационного соединения вызывающе снижается. Ионы металлов Zn (II) являются центральными атомами в их комплексах, и благодаря их меньшему размеру (0,74 А °) по сравнению с размером Cd (II) (0,97 А °) ионы металлов образуются более стабильно.
Следовательно, ион Al 3+ имеет наибольший заряд ядра, но его размер наименьший, а ион N 3- имеет наименьший заряд ядра, а его размер наибольший [17]. Инертные атомы, такие как неон, не участвуют в образовании ковалентного или ионного соединения, и эти атомы не входят в изоэлектронные ряды; следовательно, нелегко измерить радиус атомов этого типа.
Свойства стабильности зависят от размера иона металла, используемого в комплексах, и полного заряда на нем.Если размер этих ионов металлов невелик, а общий заряд велик, то их комплексы будут более стабильными. То есть их соотношение будет зависеть от заряда / радиуса. Это можно продемонстрировать с помощью следующей реакции:
Fe3 ++ 6CN − ⇆FeCN63-logβ = 31More StableE19
Fe2 ++ 6CN −⇆FeCN64-logβ = 8,3Меньше StableE20
Ионный заряд - это электрический заряд иона который формируется за счет усиления (отрицательный заряд) или потери (положительный заряд) одного или нескольких электронов из атома или группы атомов.Если говорить о стабильности координационных соединений, мы обнаружим, что общий заряд их центральных ионов металлов влияет на их стабильность, поэтому, когда мы меняем их заряд, их стабильность в диапазоне постоянных может быть определена путем распространения ошибки [18]. , Если заряд центрального металлического иона высокий, а размер небольшой, стабильность соединения высокая:
Li +> Na +> K +> Rb +> Cs + E21
Th5 +> Y3 +> Ca2 +> Na + и La3 +> Sr2 + > K + E22
В целом, наиболее стабильные координационные связи могут вызывать более мелкие и высоко заряженные рационы с образованием более стабильных координационных соединений.
Когда электронная пара притягивает к себе центральный ион, образуется сильный комплекс стабильности, что связано с донорством электронов от лиганда → иона металла. Этот процесс донорства повышает стабильность связи комплексов металлов, оказывал поляризующий эффект на некоторые ионы металлов. Li + , Na + , Mg 2+ , Ca 2+ , Al 3+ и т. Д. Представляют собой такой тип катиона металла, который не способен так сильно притягиваться из высокоэлектроотрицательного, содержащего стабильный комплексы, и эти атомы представляют собой O, N, F, Au, Hg, Ag, Pd, Pt и Pb.Лиганды такого типа, содержащие атомы P, S, As, Br и I, образуют устойчивый комплекс, поскольку они принимают электрон от M → π-связи. Ионы металлов Hg 2+ , Pb 2+ , Cd 2+ и Bi 3+ также являются электроотрицательными ионами, которые образуют нерастворимые соли сульфида металла, которые нерастворимы в водной среде.
Летучие лиганды могут быть потеряны при более высокой температуре. Это подтверждается потерей воды гидратами и аммиаком:
CoNh46Cl3∆175-180 ° C → CoNh45ClCl2 + Nh4E23
Превращение некоторых координационных соединений из одного в другое показано следующим образом:
AgHgAgI4red45 ° C⇆Ag2Hg44
Лиганд - это ион или небольшая молекула, которая связывается с атомом металла (в химии) или с биомолекулой (в биохимии), образуя комплекс, такой как координационный комплекс железо-цианид прусского голубого или железо-синий. содержащий гемоглобин белка крови. Лиганды расположены в спектрохимических рядах, которые основаны на порядке их напряженности поля. Невозможно сформировать весь ряд, изучая комплексы с одним ионом металла; ряд был разработан путем наложения различных последовательностей, полученных из спектроскопических исследований [19].Порядок общих лигандов в соответствии с их возрастающей напряженностью поля лигандов составляет
O22− Вышеуказанные спектрохимические ряды помогают нам определить прочность лигандов. Левый последний лиганд является более слабым лигандом.Эти более слабые лиганды не могут насильственно связать 3d-электрон и образовавшиеся в результате внешние октаэдрические комплексы. Это как -Mn2 + При увеличении степени окисления значение Δ увеличивается. Δ увеличивается сверху вниз. Однако, когда мы рассматриваем ион металла, наблюдаются следующие две полезные тенденции: Δ увеличивается с увеличением степени окисления. Δ увеличивается вниз по группе. Для определения константы стабильности важную роль играет природа лиганда. Следующие факторы описали природу лигандов. Размер и заряд являются двумя факторами, которые влияют на производство металлических комплексов. Меньшие заряды и небольшие размеры лигандов более благоприятны для менее стабильного образования связи с металлом и лигандом. Но если это условие как раз напротив продукта из металла и лиганда, будет более стабильное соединение.Таким образом, меньший заряд ядра и больший размер = менее стабильный комплекс, тогда как, если больший заряд ядра и маленький размер = менее стабильный комплекс. Мы берем фторид в качестве примера, потому что из-за их меньшего размера, чем у других галогенидов, и их наибольшей электроотрицательности, чем у других галогенидов, образуются более стабильные комплексы. Итак, фторид-ионные комплексы более стабильны, чем другие галогениды: FeF2 + logβ = 6.0E26 FeCl2 + logβ = 1.3E27 по сравнению с ионом S 2-, ионами O 2 2- образуются более устойчивые комплексы. Кальвин и Уилсон предполагают, что комплексы металлов будут более стабильными, если основной характер или сила лигандов выше. Это означает, что донорская способность лигандов центральным ионам металла велика [20]. Это означает, что донорская способность лигандов центральным ионам металла высока. В случае комплексообразования алифатических диаминов и ароматических диаминов стабильный комплекс образуется из алифатических диаминов, тогда как нестабильный координационный комплекс образуется с ароматическими диаминами.Итак, из вышеприведенного обсуждения мы находим, что стабильность будет выше, если сила электронного пожертвования будет больше. Таким образом, ясно, что большая базовая мощность электронодонорных частиц будет всегда формировать стабильный комплекс. NH 3 , CN - и F - вели себя как лиганды и образовывали стабильные комплексы; с другой стороны, они носят более базовый характер. Мы знаем, что если концентрация координационной группы выше, эти координационные соединения будут существовать в воде в виде раствора.Отмечено, что большую координационную тенденцию показывают молекулы воды, чем координационная группа, которая изначально присутствует. SCN - (тиоцинат) ионы присутствуют в более высокой концентрации; с ионом металла Co 2+ он образовал комплекс синего цвета, который стабилен в состоянии, но при разбавлении водной среды вместо синего образуется розовый цвет, или синий комплекс разрушается [Co ( H 2 O) 6 ] 2+ , и теперь, если мы добавим еще SCN - , розовый цвет не появится: CoSCN42− + h3O⇆Coh3O62 ++ 4SCN − BluePinkE28 Теперь это Ясно, что H 2900 O и SCN - конкурируют за образование металлсодержащего комплексного соединения Co (II).В случае тетрааминового комплекса сульфата меди с медью аммиак действует как донорный атом или лиганд. Если концентрация NH 3 ниже в реакции, образуется гидроксид меди, но при более высокой концентрации образуется тетраамин сульфат меди, как в следующей реакции: CuSO4 + Nh5OH → CuOh3Малое количество лигандаE29 CuSO4 + Nh5OH → CuOh3CuNh52SO4.h3OHвысокая концентрация лигандаE30 Для иона металла хелатный лиганд усиливается и сродство, и это известно как хелатный эффект и сравнивается с не хелатирующим и монодентатным лигандом или мультидентатным лигандом, действующим как хелатирующий лиганд агент.Этилендиамин является простым хелатирующим агентом (рис. 1). Структура этилендиамина. Из-за бидентатной природы этилендиамина он образует две связи с ионом металла или центральным атомом. Вода образует комплекс с ионом металла Ni (II), но благодаря своей монодентатной природе она не является хелатирующим лигандом (рис. 2 и 3). Структура хелатной конфигурации этилендиаминного лиганда. Структура хелата с тремя этилендиаминными лигандами. Дентатный мошенник лиганда обеспечивает прочность связи с ионом металла или центральным атомом, и по мере увеличения количества дентата герметичность также увеличивается. Это явление известно как хелатирующий эффект, в то время как образование комплексов металлов с этими хелатирующими лигандами называется хелатированием: Металл + 2Лиганд-Металл-лиганд2K = ML2ML2E31 Металл + лиганд-лиганд-Метал-лиганд-лиганд E32 или Размеры хелатного кольца увеличиваются, а стабильность металлического комплекса уменьшается. Согласно Шварценбаху, соединительные мосты образуют хелатообразующие кольца. Удлиненное кольцо преобладает, когда длинные мостики соединяются с лигандом, образуя длинное кольцо. Обычно наблюдается, что увеличение размера хелатного кольца приводит к снижению стабильности комплекса. Он истолковал это утверждение. Энтропия комплекса будет изменяться, если увеличится размер хелатного кольца, т.е.второй донорный атом разрешен хелатирующим кольцом. По мере увеличения размера хелатирующего кольца стабильность должна увеличиваться с эффектом энтропии. Четырехчленные кольцевые соединения являются нестабильными, тогда как пятичленные являются более стабильными. Таким образом, хелатообразующее кольцо увеличило его размер и стабильность образующихся металлических комплексов. Количество хелатных колец также определяет стабильность комплексов. Не хелатообразующие соединения металлов менее стабильны, чем хелатообразующие соединения.Эти числа увеличивают термодинамический объем, и это также известно как термин энтропии. В последние годы были описаны лиганды, способные занимать до шести координационных положений на одном ионе металла. Сообщалось об исследованиях констант образования координационных соединений с этими лигандами. Количество лигандов или хелатообразующих агентов влияет на стабильность комплексов металлов, поэтому, когда эти числа увеличиваются и уменьшаются, стабильность также будет изменяться вместе с ним. Для комплексов Ni (II) с этилендиамином в качестве хелатирующего агента его log K 1 составляет 7.9, и если хелатирующими агентами являются трин и пентен, то значения log K 1 составляют 7,9 и 19,3 соответственно. Если вместо Ni (II) используется замена иона металла Zn, то значения log K 1 для этилендиамина, трина и пентена составляют 6,0, 12,1 и 16,2 соответственно. Значения logMMY для ионов металлов приведены в таблице 1. Значения ионов металлов против log βMY. Ni (NH 3 ) 6 2+ представляет собой октаэдрический комплекс металлов, и при 25 ° C его log β 6 составляет 8,3, но Ni (этилендиамин) 3 2+ комплекс также является восьмигранным в геометрии, с 18,4 в качестве значения log β 6 .Рассчитанное значение стабильности Ni (этилендиамина) в 3 2+ 10 10 раз является более стабильным, поскольку этилендиамин образует три кольца в виде хелатообразующих колец по сравнению с тем, что такое кольцо не образуется. Этилендиаминтетраацетат (ЭДТА) представляет собой гексадентатный лиганд, который обычно образует стабильные комплексы металлов благодаря своей хелатирующей способности. Специальный эффект в молекулах - это когда атомы занимают пространство. Это называется стерическим эффектом. Энергия необходима для сближения этих атомов друг с другом.Эти электроны убегают от ближних атомов. Там может быть много способов его создания. Мы знаем отталкивание между валентными электронами как стерический эффект, который увеличивает энергию системы тока [21]. Благоприятный или неблагоприятный любой ответ создан. Например, если статический эффект больше, чем у продукта в процессе образования металлокомплекса, то статическое увеличение будет способствовать этой реакции. Но если дело обстоит иначе, скептицизм будет направлен на отсталость. Этот эффект будет в основном зависеть от конформационных состояний, и теория минимального стерического взаимодействия также может быть рассмотрена.Эффект вторичной стерики виден на связывание рецептора, производимое альтернативой, такой как: Ограниченный доступ к критической группе. Палка-барьер. Электронная резонансная замещающая связь путем отталкивания. Население конформера изменяется из-за эффекта активного экранирования. Макроциклический эффект аналогичен изображению хелатного эффекта.Это означает, что принцип обоих одинаков. Но макроциклический эффект предполагает циклическую деформацию лиганда. Макроциклические лиганды более испорчены, чем хелатирующие агенты. Скорее, их соединения более стабильны из-за их циклически ограниченного сужения. Требуется некоторая энтропия в организме для реакции с ионом металла. Например, для тетрадентатного циклического лиганда мы можем использовать гем-B, который образует комплекс металлов с использованием ионов Fe +2 в биологических системах (рис. 4). Структура гемоглобина является биологическим комплексным соединением, которое содержит ион металла Fe (II). Н-дентатные хелатообразующие агенты играют важную роль для образования более стабильных комплексов металлов по сравнению с н-унденатными лигандами. Но н-дентатный макроциклический лиганд дает более стабильную среду в комплексах металлов по сравнению с лигандами с открытой цепью. Это изменение очень благоприятно для изменения энтропии (ΔS) и энтальпии (ΔH). Печень - это орган, который выполняет множество важных функций в организме. Однако, если вы наблюдаете его под микроскопом, кажется, что он очень тихий, потому что большая часть его клеток остается в неделящемся состоянии (это называется покоем). Печень настолько важна, что обладает большой способностью переносить несколько видов травм. После раны или другого повреждения все клетки печени изменяются и делятся, пока не восстановится нормальный размер печени.Этот интересный процесс широко известен как «регенерация печени». Если повреждение печени настолько серьезное, что печень не может регенерировать, человеку может понадобиться новая печень, то есть пересадка. Сейчас более 15 000 человек в США ждут пересадки печени. Вы узнаете, что благодаря своей способности «регенерировать» можно пожертвовать печень, пока донор еще жив. Печень - самый большой орган, расположенный в брюшной полости; весит около 1.5 кг у человека и организована в секции, называемые долями [1]. Печень похожа на большую фабрику, где некоторые молекулы производятся, а другие разрушаются, чтобы получить энергию, необходимую организму для работы. Печень может накапливать энергию и использовать ее во время длительного голодания или во время сна. Печень может избавляться от токсичных веществ в организме, таких как алкоголь, лекарства или наркотики, путем производства вещества, называемого желчью (этот процесс называется детоксикация ). Желчь - это вид сока, который действует как детергент, расщепляя определенные вещества и переваривая жиры.Поскольку невозможно смешать воду и масло, печень использует желчь, чтобы поймать капли жира в «пузыре». Структура пузырьков позволяет им путешествовать по водной среде тела. После выработки печенью желчь транспортируется через небольшие сосуды (желчные канальцы), расположенные в гепатоцитах, в соседний орган, называемый желчным пузырем, затем в кишечник, где восстанавливаются питательные вещества, и затем возвращается в печень. Маршрут желчи следует из печени в кишечник и известен как желчное дерево .Кроме того, печень действует как часть иммунной системы, так как помогает организму бороться с инфекционными организмами. Для выполнения всех этих сложных задач печень должна содержать несколько типов клеток (рисунок 1). На первый взгляд печень имеет глубокий красный цвет. Это потому, что он содержит множество кровеносных сосудов или синусоид, транспортирующих кровь из кишечника, селезенки и поджелудочной железы. Наиболее многочисленными и самыми большими клетками печени являются гепатоцитов , которые являются клетками, которые производят и разрушают определенные молекулы, чтобы дать организму энергию.Другие клетки помогают печени работать. Клетки Купфера - это клетки-хранители печени, которые питаются чужеродными бактериями. Звездообразные печеночных звездчатых клеток хранят витамин А, который выглядит как яркие капли внутри этих клеток. Звездчатые клетки печени также помогают продуцировать белки, которые составляют структуру печени. синусоидальных эндотелиальных клеток покрывают обильные кровеносные сосуды печени. На данный момент, я надеюсь, вы понимаете, что печень является важным органом; никто не может выжить без него.Печень имеет интересные механизмы, чтобы защитить себя от различных травм и болезней. В нормальных условиях мы говорим, что печень является покоящимся органом . Это означает, что если вы наблюдаете это под микроскопом, вы можете найти только одну клетку, разделяющуюся на тысячи клеток! Однако, если печень физически повреждена (например, рана) или химически повреждена (наркотиками или алкоголем), ее клетки выталкиваются, чтобы делиться и расти. Это явление известно как , регенерация печени , и оно даже упоминается в греческой мифологии.Самый популярный миф, который ссылается на это явление, - это Прометей, который украл огонь. Его наказание заключалось в том, чтобы оставаться в цепях в горах Кавказа, где орел каждый день питался печенью. Хотя люди давно знали о регенерации печени, только в 1900-х годах регенерация печени была официально изучена и обнаружена у всех млекопитающих. Чтобы изучить регенерацию печени, в 1931 г. Хиггинс и Андерсон провели эксперимент на крысах, в котором две самые большие доли печени были очень тщательно удалены [2].Работа с животными предполагает использование чистых инструментов, анестезию и бережное отношение. Поскольку в печени много кровеносных сосудов, чтобы избежать кровотечения, основные сосуды печени были перевязаны специальной нитью. Хиггинс и Андерсон обнаружили, что печень крысы вернулась к нормальному размеру за 7 дней. Даже сегодня это самый популярный для ученых способ изучения регенерации печени. Используя эксперименты такого типа, мы можем понять, какие гены, гормоны и белки факторов роста участвуют в процессе регенерации печени. Несмотря на то, что мы называем повторный рост печени «регенерацией печени», это не совсем точное описание того, что происходит. Термин «регенерация» фактически описывает процесс, посредством которого животные могут заменить целую часть тела. Например, если ящерица теряет свой хвост, убегая от хищника, его хвост может в конечном итоге отрастать. Этого не может случиться с печенью - если кто-то потеряет свою печень целиком, он или она не сможет просто вырастить новую.Таким образом, хотя мы будем продолжать называть это «регенерацией печени», более точный термин для того, что происходит в печени, - это компенсаторная гиперплазия [3]. Хотя этот термин может показаться сложным, на самом деле он означает увеличение размера печени, вызванное увеличением количества клеток. Чтобы иметь возможность делиться, клетки печени должны «проснуться» или, как говорят ученые, клетки должны стать «активированными». Процесс регенерации печени происходит в три этапа (рис. 2). На трех этапах все ячейки работают как команда и общаются друг с другом.Это общение говорит клеткам печени выйти из состояния покоя и начать делиться. Через несколько минут после того, как печень обнаруживает повреждение, гены в ядрах клеток печени посылают сигналы для подготовки клеток к делению. Это называется фазой инициирования . Во время фазы пролиферации клетки печени делятся. Деление клеток продолжается до тех пор, пока печень не восстановит свой первоначальный размер, затем наступает фаза завершения и , и клетки печени прекращают делиться. Пока все это происходит, печень все еще должна выполнять все свои нормальные функции, чтобы животное оставалось живым и здоровым. Как печень узнает, когда перестать расти? Это важный вопрос, и мы можем сказать, что размер печени является очень важным фактором. Печень составляет около 3% тела, и эта пропорция имеет тенденцию оставаться постоянной, независимо от размера тела.Если вы хотите узнать размер своей печени, просто умножьте свой вес на 0,03! Такая же пропорция применяется у других млекопитающих. Таким образом, если маленькая собака получает печень большой собаки, печень будет расти в соответствии с размером тела маленькой собаки. Некоторые ученые дали название «гепатостат» (например, «термостат», только «гепато» означает печень) для системы, которая регулирует размер печени. В общем, мы можем представить, что гепатостат содержит набор белков, которые посылают сигналы остановки в печень, так что он больше не будет расти.Обратите внимание, что эти «стоповые» белки имеют противоположные эффекты от тех, которые действуют на фазе инициации (те, которые дают приказ «делиться»). Ученым все еще нужно проделать большую работу, чтобы понять, как контролируется гепатостат и факторы, которые на него влияют. Многие знания, полученные учеными, могут быть использованы для решения конкретных задач в краткосрочной или долгосрочной перспективе. Поскольку регенерация печени включает хорошо контролируемый рост, мы можем использовать наши знания о регенерации печени, чтобы помочь нам понять другие типы роста, такие как неконтролируемый рост, который происходит в раковых клетках. Врачи могут использовать знания о регенерации печени, чтобы помочь людям с заболеваниями или повреждениями печени, которые выходят за пределы ее способности к регенерации. В крайних случаях этим людям может понадобиться пересадка печени. Полные печени или части печени могут быть получены от мертвых доноров и пересажены пациентам. Но теперь, когда вы знаете о регенеративной способности печени, вы можете понять, почему здоровый, живой человек также может пожертвовать кусочек своей печени кому-то, кто нуждается в пересадке.Печень донора отрастет! Хотя этот тип трансплантации от живого донора возможен, доноров недостаточно для того, чтобы справиться с большим количеством людей, которым необходима пересадка печени. В настоящее время в США почти 15 000 человек нуждаются в новых печенках [4]. Кроме того, ученые работают над созданием устройств, чтобы помочь пациентам с определенными проблемами печени. Эти устройства могут временно помогать пациентам в ожидании пересадки, аналогично диализным аппаратам, которые используются для помощи пациентам с почечной недостаточностью [5]. Тканевая инженерия - это отрасль науки, которая также может помочь пациентам, ожидающим пересадки. Тканевая инженерия использует специальные методы и материалы, чтобы попытаться вырастить печень в лаборатории. В отсутствие доноров эти выращенные в лаборатории печени можно пересадить пациентам, чтобы улучшить или заменить их поврежденные ткани печени. Огромная часть знаний о биологическом поведении клеток печени была получена при изучении феномена регенерации печени. Принимая во внимание эти знания и тот факт, что доноров печени мало, другой стратегией тканевой инженерии является «переработка» всей печени или ее небольшой части.Можно использовать печень мертвого донора, но все исходные клетки удаляют, пропуская специальный раствор через сосуды печени, пока он не станет полностью прозрачным (рис. 3). Этот процесс называется decellularization , и это происходит потому, что трехмерная структура или матрица, которая поддерживает клеточные компоненты печени, хорошо сохраняется. Обратите внимание на хорошо сохранившиеся кровеносные сосуды матрицы печени. Таким образом, чистая печень может получать новые печеночные клетки от здоровых людей или животных и затем пересаживаться [6].Этот процесс был успешно проведен у нескольких видов млекопитающих, но это сложно. Очистка печени крысы может занять 48 часов, а чистка печени человека - до 6 недель! Однако проблема, которую необходимо решить после децеллюляризации, заключается в получении достаточного количества здоровых клеток печени для пополнения чистой печени. В настоящее время изучаемой альтернативой является производство клеток печени из стволовых клеток, расположенных в печени или из нее. Но это уже другая история. Целью написания этой статьи было рассказать молодым людям о том, как печень работает в организме, и описать ее удивительную способность реагировать на различные травмы. Другой целью было предоставить знания о научных исследованиях, направленных на улучшение качества жизни людей с тяжелыми заболеваниями печени.Я надеюсь, что благодаря этой информации вы поймете важную роль, которую ваша печень играет в здоровье вашего тела, и важность заботы о вашей печени на протяжении всей жизни. Детоксикация : ↑ Обработка токсичных веществ для их удаления из организма. Билиарное дерево : ↑ Набор сосудов, по которым желчь перемещается из печени в желчный пузырь. Quiescent : ↑ Стадия, где клетки не делятся. Компенсаторная гиперплазия : ↑ Процесс, при котором печень после повреждения восстанавливает свой размер, увеличивая количество клеток печени. Decellularization : ↑ Процесс, выполняемый для удаления клеток ткани / органа с целью сохранения матрицы, которую можно пополнить здоровыми клетками. Автор заявляет, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов. Автор хотел бы поблагодарить д-ра Алехандро Сото-Гутьерреса, который работает в Университете Питтсбурга, штат Пенсильвания, США, за то, что он поделился фотографией печеночной ткани, и г-на Патрика Вейля, который помогал с редактированием на английском языке. [1] ↑ Цунг А. и Геллер Д. А. 2011. Общая и клеточная анатомия печени. В г. Молекулярная патология заболеваний печени . редактор С.П.С. Монга. Питсбург, Пенсильвания: Springer Science + Business Media, LLC. п. 3-6. [2] ↑ Хиггинс Г. М. и Андерсон Р. М. 1931. Экспериментальная патология печени. I. Восстановление печени белой крысы после частичного хирургического удаления. Arch. Pathol. 12: 186–202. [3] ↑ Columbano, A., Shinosuka, H. 1996. Регенерация печени в сравнении с прямой гиперплазией. FASEB J. 10: 1118–28. [4] ↑ Сайт U.S. Департамент здравоохранения и социальных служб. Сеть закупок и трансплантации органов (OPTN-HRSA) . Доступно по адресу: http://optn.transplant.hrsa.gov (по состоянию на 14 марта 2017 г.). [5] ↑ Ли, С. Я., Ким, Х. Дж. И Чой, Д. 2015. Источники клеток, системы поддержки печени и инженерия тканей печени: альтернативы трансплантации печени. Int. J. Стволовые клетки 8: 36–47. doi: 10.15283 / ijsc.2015.8.1.36 [6] ↑ Уйгун, Б. Э., Soto-Gutierrez, A., Yagi, H., Izamis, M.L., Guzzardi, M.A., Shulman, C., et al. 2010. Реинжиниринг органов путем разработки трансплантируемого транспеллеризованного трансплантата печени с использованием децеллюляризованного матрикса печени. Nat. Med. 16: 814–20. doi: 10.1038 / nm.2170 Через несколько лет после идентификации и характеристики отдельных членов пути Hippo у Drosophila и млекопитающих важность передачи сигналов Hippo в печени стала очевидной благодаря поразительному фенотипу: избыточная экспрессия YAP или экспрессия активированного YAP приводила к значительному чрезмерному росту печени идентифицируя передачу сигналов Hippo как важную детерминанту в контроле размера органа (Camargo et al.2007; Dong et al., 2007). Быстрое развитие гепатоцеллюлярной карциномы (ГЦК) при избыточной экспрессии YAP дополнительно подтвердило мощную онкогенную роль этого белка (Dong et al., 2007). Недавно исследование передачи сигналов Hippo в непаренхимных клетках печени, включая печеночные звездчатые клетки (HSC) и синусоидальные эндотелиальные клетки печени (LSEC), дало представление о сложном взаимодействии между различными типами печеночных клеток с глубоким влиянием на патофизиологию печени. болезнь. Здесь мы даем обзор сигналов Hippo в печени, включая последние достижения и открытые вопросы, а также будущие направления в этой области. Вскоре после открытия функции YAP в мышиной печени протеинкиназы MST1 и MST2 были подтверждены в качестве регуляторов пути Hippo вверх по течению, которые ограничивают активацию YAP, избыточный рост тканей и канцерогенез (Figure 1; Zhou et al., 2009; Lu et al., 2010; Song et al., 2010). В той же линии печеночная инактивация MST1 / 2-адапторного белка SAV1 / WW45 привела к пролиферации клеток, ассоциированной с YAP, и у мышей-мутантов в конечном итоге развились опухоли с характеристиками ГЦК и внутрипеченочных холангиокарцином (ICC) (Lee et al.2010; Lu et al., 2010). Условный нокаут Nf2 , гомолога млекопитающего вышестоящего регулятора Hippo Merlin , привел к снижению фосфорилирования Lats1 / 2 и, тем самым, активации YAP, что привело к чрезмерному росту печени и развитию опухолей печени (Benhamouche et al. 2010; Zhang et al., 2010). Важно отметить, что эти данные установили, что NF2 является негативным регулятором YAP, и подтвердили, что большая часть пути Hippo Drosophila сохраняется у млекопитающих (Figure 1). Рисунок 1. Ключевые компоненты пути Бегемота у дрозофилы и млекопитающих. Во всех этих моделях условная инактивация генов пути Hippo была достигнута с использованием трансгенных мышей CAGGCre-ER (Zhou et al., 2009; Song et al., 2010) или альбумина -Creed Cre . (Benhamouche et al., 2010; Lu et al., 2010; Zhang et al., 2010), который также активен в фетальных гепатобластах, которые дают клетки желчных протоков.Мыши-мутанты демонстрировали различную степень пролиферации гепатоцитов, но также демонстрировали пролиферацию и расширение популяции клеток печени с небольшими ядрами вокруг портальной триады, так называемые «овальные клетки». Долгое время считалось, что эти клетки функционируют как бипотентные клетки-предшественники печени, которые могут дифференцироваться в гепатоциты и клетки желчных протоков при определенных условиях, таких как серьезное повреждение гепатоцитов - гипотеза, которая была оспорена недавними исследованиями (Tanimizu and Mitaka, 2014). Экспансия овальных клеток и развитие как HCC, так и ICC первоначально привели к предположению, что опухоли в моделях, инактивированных путем Hippo, возникают из этих потенциальных бипотентных клеток-предшественников.Однако недавние исследования показывают, что эти фенотипы возникают в результате трансдифференцировки мутантных гепатоцитов и дерегулированного морфогенеза желчевыводящих путей (Yimlamai et al., 2014; Benhamouche-Trouillet et al., 2018). Несколько гепатоцит-специфических моделей трансфекции могут инициировать развитие опухолей со смешанной дифференцировкой: сверхэкспрессия YAP, а также инактивация вышестоящего регулятора Hippo Nf2, опосредованного AAV-Cre, индуцирует дедифференцировку гепатоцитов к фенотипу, подобному предшественнику (Yimlamai et al. ,2014). Кроме того, гидродинамическая инъекция в хвостовую вену экспрессирующих конструкций на основе транспозона для конститутивно активных YAP и PIK3CA - каталитической субъединицы PI3K - приводила к образованию опухолей печени с гепатоцеллюлярной, холангиоцеллюлярной или смешанной дифференцировкой HCC / ICC. В этой модели опухоли характеризовались активацией путей mTORC1 / 2, ERK / MAPK и Notch (Li et al., 2015). На сегодняшний день молекулярная основа для сотрудничества между передачей сигналов PI3K и YAP при раке печени не совсем понятна, но может быть опосредована индуцированной PI3K активацией CD166, белка клеточной поверхности, который, как было показано, положительно регулирует активность YAP (Ma et al. и др.2014). С другой стороны, данные эпителиальных клеток молочной железы и клеток рака толстой кишки указывают на то, что передача сигналов PI3K / PDK1 / AKT способствует активности YAP через LATS-зависимые и -независимые механизмы (Zhao et al., 2018). Однако, если этот механизм сохраняется при раке печени и как он связан с клеточной дифференцировкой, еще предстоит выяснить. Из того, что известно на сегодняшний день, YAP - и, возможно, другие онкогенные пути, такие как передача сигналов PI3K - не только способствуют пролиферации и онкогенезу в целом, но и онкогенной пластичности гепатоцитов с трансдифференцировкой к фенотипу, похожему на предшественник или даже к желчному фенотипу ( Yimlamai et al.2014; Fitamant et al., 2015; Font-Burgada et al., 2015; Патель и др., 2017). Сигнализация Notch - критический путь в развитии желчных протоков - является ключевым кандидатом для запуска этого дифференцирующего переключателя. Следует отметить, что передача сигналов Notch может быть активирована посредством активации генов-мишеней YAP, таких как Notch3 и Jag1 (Jeliazkova et al., 2013; Tschaharganeh et al., 2013; Yimlamai et al., 2014; Wu et al. , 2017; Zhang S. et al., 2018). Однако исследование с использованием печеночно-специфического нокаута киназ LATS1 и LATS2 пути Hippo не подтвердило роль передачи сигналов Notch, но показало, что активная передача сигналов TGFβ ниже YAP индуцирует транс-дифференцировку в клетки желчных протоков (Lee et al., 2016). Кроме того, YAP способствует связыванию регуляторов транскрипции HNF4A и FOXA2 с сайтами эмбриональных энхансеров во время дифференцировки гепатоцитов, чтобы усилить транскрипцию эмбрион-специфических генов - механизм, который может быть угнан в трансформированных клетках для стимулирования дедифференцировки и онкогенеза (Alder et al., 2014) , Взятые вместе, сложный паттерн взаимодействий онкогенного пути определяет пластичность гепатоцитов и дифференцировку опухоли при инактивации пути Бегемота, что мы только начинаем понимать. Помимо стимуляции дифференцировки желчных путей в гепатоцитах, передача сигналов YAP имеет большое значение для клеток желчных протоков. Высокая цитоплазматическая и ядерная экспрессия YAP в холангиоцитах у человека и мышей печени позволяет предположить, что активность YAP имеет отношение к клеткам желчных протоков, но функциональные исследования, анализирующие роль передачи сигналов Hippo в холангиоцитах, ограничены. (Bai et al., 2012; Patel et al., 2017). Тем не менее было сделано несколько интересных наблюдений в на основе Albumin-Cre моделей условного нокаута, которые активируют Cre в предшественниках как гепатоцитов, так и холангиоцитов (Geisler et al. ,, 2008): условный нокаут одного YAP или как YAP, так и его ортопеда TAZ - также известного как WWTR1 - приводит к дефектам морфогенеза желчных протоков с неправильной формой и деформацией внутрипеченочных желчных протоков (Zhang et al., 2010; Lu et al. al., 2018), в то время как нокаут Nf2 приводил к расширению билиарных структур (Benhamouche-Trouillet et al., 2018). У людей YAP активируется при протоковых реакциях при холестатической болезни печени и при неалкогольном стеатогепатите (Bai et al.2012; Anakk et al., 2013; Machado et al., 2015). При холестатическом повреждении печени после перевязки желчных протоков у мышей эта реактивная пролиферация и расширение клеток желчных протоков заметно снижается при недостаточности YAP в печени, что указывает на то, что активность YAP необходима для формирования протоковых реакций. У взрослых мышей генетическая инактивация YAP с использованием индуцибельной системы Mx-Cre приводит к глубокому повреждению гепатоцитов и уменьшению компенсаторной пролиферации гепатоцитов и клеток желчных протоков, что связано с повышенной смертностью по сравнению с мышами дикого типа (Bai et al.2012). Механистически, накопление желчных кислот при холестатическом заболевании печени, по-видимому, является важным активатором YAP ниже по течению белка белка IQGAP1, который индуцируется желчными кислотами - механизм, который также может играть роль в канцерогенезе (Anakk et al., 2013) , Механистически, существует немало свидетельств того, что IQGAP1 может напрямую связываться с YAP, чтобы изменять его транскрипционную активность (Sayedyahossein et al., 2016). Однако это взаимодействие, по-видимому, является в основном тормозящим, что указывает на то, что другой, пока неизвестный механизм может быть ответственен за IQGAP1-опосредованную активацию YAP в печени.При холангиокарциноме человека активированный YAP ассоциируется с плохим прогнозом, химической стойкостью, ангиогенезом и хромосомной нестабильностью (Marti et al., 2015; Wu et al., 2016; Rizvi et al., 2018), а высокая экспрессия TAZ коррелирует с уменьшением выживаемость после резекции опухоли (Xiao et al., 2016). У мышей экспрессия активированных YAP и AKT в гепатоцитах является достаточной для индукции холангиокарциномы, зависящей от передачи сигналов AKT / mTOR, которая, вероятно, взаимодействует с YAP в онкогенезе и, возможно, способствует активности YAP (Zhang et al., 2017; Чжао и др., 2018). Кроме того, YAP важен для поддержания фенотипа холангиоцитов, так как ингибирование YAP путем сверхэкспрессии либо LATS2, либо доминантно-негативного варианта TEAD2 приводило к снижению экспрессии маркеров холангиоцитов (Zhang S. et al., 2018). Долгое время оставалось загадкой, как восстановление размера печени почти точно до ее первоначального объема надежно достигается после резекции печени.Передача сигналов гиппопотама, казалось, была идеальным кандидатом для пути, который управляет контролируемой активацией и прекращением пролиферации гепатоцитов во время регенерации. Действительно, существует активная активация YAP / TAZ ниже по течению примерно через 24 ч после частичной гепатэктомии с последующей активацией ингибирующих киназ Hippo, возможно, для ограничения чрезмерной активации YAP / TAZ и остановки пролиферации, когда печень приближается к своему первоначальному объему (Grijalva et al., 2014; Loforese et al., 2017; Lu et al., 2018). Специфичный для печени нокаут YAP и TAZ приводит к нарушению регенерации печени и задерживает восстановление массы печени (Lu et al., 2018), подтверждая важность передачи сигналов Hippo в регенерации печени. Напротив, YAP и TAZ не являются обязательными для завершения процесса регенерации, поскольку альтернативные пути могут заменить их активность для обеспечения регенерации печени, хотя и с существенной задержкой (Michalopoulos, 2017). В развитии плода опосредованный Alb-Cre условный нокаут YAP в гепатоцитах и холангиоцитах, начиная с эмбрионального дня 13,5, приводил главным образом к дефектному развитию внутрипеченочных желчных протоков - но интересно, что явного фенотипа в гепатоцитах не было до 8 недель после рождения (Zhang et al. и др.2010). Частично это открытие было объяснено спасением функции YAP за счет повышенной активности его ортолога TAZ, сходного с фенотипами, наблюдаемыми у рыбок данио (Yi et al., 2018). Но что поразительно, удаление как YAP, так и TAZ с помощью Albumin-Cre приводит к развитию почти нормальной печени - и, как это ни парадоксально, приводит к увеличению размера печени. Этот неожиданный фенотип может быть объяснен увеличением повреждения гепатоцитов с последующей активацией других путей пролиферации, которые неспособны контролировать печеночную массу так же жестко, как передача сигналов Hippo (Zhang et al.2010; Lu et al., 2018). Эти данные подчеркивают, что активация и взаимодействие пути в эмбриональном развитии и во время регенерации печени характеризуется высокой пластичностью, чтобы приспособиться к функциональному дефициту основных пролиферативных путей, таких как передача сигналов YAP / TAZ. Однако эта функциональная избыточность, по-видимому, не работает так эффективно, как только мыши достигают определенного возраста: пожилые или больные печени часто не могут в достаточной степени регенерировать, поскольку они не способны поддерживать адекватную пролиферативную передачу сигналов.В некоторой степени этот дефект регенерации обусловлен гиперактивной передачей сигналов Hippo - которая не в достаточной мере устранена компенсаторной активацией других пролиферативных путей по причинам, которые до сих пор не совсем понятны. Глушение MST-киназ с помощью инкапсулированной в липосомы миРНК восстанавливало экспрессию генов-мишеней YAP и улучшало регенерацию печени у пожилых животных (Loforese et al., 2017). В другом исследовании новый ингибитор MST2 уменьшал апоптоз и повреждение печени при острой печеночной недостаточности, вызванной аминоацетофеном.Поразительно, что аналогичный эффект, включая стихание фиброза, наблюдался при хроническом повреждении печени после перевязки желчных протоков или повторного введения CCl и 4 954 соответственно (Fan et al., 2016). Ориентация вверх по течению гиппинкиназ при острой или хронической печеночной недостаточности у человека для восстановления регенерации является многообещающим подходом с высокой клинической значимостью, но его необходимо сопоставить с риском онкогенной трансформации, особенно при цирротической печени, которая по своей природе имеет высокий риск развития ГЦК. Звездчатые клетки печени являются основным типом клеток, вовлеченных в фиброз печени, но также играют критическую роль в остром восстановлении после повреждения печени или частичной гепатэктомии (Kordes et al., 2014). Хроническое повреждение печени запускает постоянную активацию звездчатых клеток с секрецией и накоплением избыточных белков внеклеточного матрикса (ECM), транс-дифференцировкой спокойных HSCs в миофибробласты, накапливаясь в образовании фиброзной рубцовой ткани.Результат этого нерегулируемого процесса восстановления продиктован молекулярными факторами, которые контролируют активацию HSC, которые поэтому рассматриваются как кандидаты на целевую терапию. Недавно передача сигналов Hippo была идентифицирована как важный путь активации звездчатых клеток. YAP активируется в HSCs во время острой регенерации печени после частичной гепатэктомии или ишемии-реперфузионного повреждения и, соответственно, при хроническом повреждении фиброзной печени человека, а также при индуцированном CCl 4 фиброзе у мышей (Mannaerts et al.2015; Swiderska-Syn и др., 2016; Konishi et al., 2018). Важно, что устойчивая активация YAP при фиброзе печени связана с увеличением белков ЕСМ и жесткости тканей (Dechene et al., 2010). Влияние жесткости матрикса на передачу сигналов YAP хорошо установлено, и механические сигналы все чаще признаются в качестве медиатора патологической активации YAP in vivo : повышенная жесткость матрикса связана с активацией рецепторов клеточной поверхности, которые регулируют полимеризацию F-актина в RhoA -зависимый способ стимулирования ядерной транслокации и активации YAP / TAZ - вероятно, обеспечивает молекулярную основу для активации YAP в HSC и гепатоцитах при фиброзе и, возможно, усиливает канцерогенез в цирротических печенках (Dupont et al.2011; Жубанчалиев и др., 2016). При остром повреждении печени другие пути, такие как передача сигналов Hedgehog, вероятно, способствуют активации YAP. После частичной гепатэктомии мыши с HSC, дефицитными по функциональной передаче сигналов Hedgehog, обнаруживают нарушенную активацию YAP в звездчатых клетках, а также в гепатоцитах (Swiderska-Syn et al., 2016), что указывает на то, что передача сигналов Hedgehog / YAP обеспечивает важную перекрестную связь между этими клетками типы для обеспечения достаточной регенерации. В то время как активность HSC важна для целостности печени после острого повреждения, устойчивая активация вредна для хронического повреждения печени, что делает YAP потенциальной мишенью для предотвращения активации HSC и прогрессирования фиброза.Важно отметить, что фармакологическое ингибирование YAP / TEAD вертепорфином заметно снижает активацию HSC и затрудняет фиброгенез, который обычно происходит после лечения CCl 4 у мышей (Mannaerts et al., 2015; Martin et al., 2016). Другими непаренхиматозными клетками, играющими существенную роль в повреждении печени, являются синусоидальные эндотелиальные клетки печени (LSEC), которые, как было показано, влияют на регенерацию и фиброз через ангиокринную передачу сигналов звездчатым клеткам при остром и хроническом повреждении печени (Ding et al.2014; Пуассон и др., 2017). Эксперименты по совместному культивированию показали, что дифференцированные LSECs поддерживают HSC покоя и что нарушение регуляции перекрестных помех между гепатоцитами, LSECs и HSCs при хроническом повреждении печени способствует фиброзу (Poisson et al., 2017). Интересно, что LSEC также важны для поддержания целостности клеток после активации YAP в отдельных гепатоцитах. Комбинированное повреждение гепатоцитов и LSEC, вызванное этанолом, но не повреждение только гепатоцитов или LSEC, меняет судьбу активированных YAP гепатоцитов от пролиферации до апоптоза (Miyamura et al., 2017). Было показано, что активация YAP в LSEC регулирует экспрессию белков HIF-1α и VEGF-A, способствуя ангиогенезу при фиброзе - процессе, блокирующем транспорт питательных веществ из синусоид и усиливающем повреждение печени (Elpek, 2015; Zhang C. et al., 2018 ). Помимо запуска неоангиогенеза в больной печени, весьма вероятно, что YAP / TAZ также играют важную роль в формировании печеночных кровеносных сосудов во время развития печени, в том числе в прорастании эндотелиальных клеток (EC) и созревании соединения (Park and Kwon, 2018 ).Тем не менее, соответствующая экспрессия YAP / TAZ не может быть обнаружена в печеночных ECs в P5 и P12, что в отличие от эндотелиальных клеток в развивающейся сетчатке и мозге в те же моменты времени - ткани, где YAP и TAZ функционально необходимы для правильного ангиогенеза (Kim et al., 2017a). Хотя это может указывать на то, что передача сигналов YAP / TAZ может не иметь отношения к ангиогенезу печени, более вероятно, что активация YAP и TAZ действительно происходит в более ранние моменты времени во время развития печени, где формирование EC начинается уже в E9.5 (Чжао и Дункан, 2005). Если активация передачи сигналов YAP / TAZ в эндотелиальных клетках играет значительную роль при фиброзе и других заболеваниях печени, тем не менее, это еще предстоит изучить. В связи с растущей важностью передачи сигналов Hippo в HSC, LSECs и, возможно, эндотелиальных клетках при фиброзе и повреждениях, любые соображения, направленные на терапевтическую направленность этого пути, должны учитывать, что регенерация печени, вероятно, будет нарушена - возможно, усугубит повреждение печени в долгосрочной перспективе. В любой ткани пролиферация и рост клеток зависят от доступности питательных веществ.В этом контексте вероятна регуляция пролиферативного YAP и TAZ посредством клеточного метаболизма. Действительно, низкие уровни энергии запускают ингибирование YAP и TAZ, в то время как высокие уровни глюкозы и жирных кислот активируют транскрипцию YAP / TAZ вниз по течению. С другой стороны, было показано, что сами YAP / TAZ способствуют ключевым метаболическим процессам, таким как гликолиз, для обеспечения адекватного снабжения энергией пролиферирующих клеток (Koo and Guan, 2018). В печени избыточная доступность питательных веществ может привести к стеатозу и стеатогепатиту (неалкогольная жировая болезнь печени или НАЖБП), при которой накопление жира приводит к воспалению и разрушению гепатоцитов, способствуя развитию цирроза и рака печени. На мышиных моделях НАЖБП и в образцах стеатогепатита человека может наблюдаться повышение уровня YAP / TAZ - главным образом при регенеративных протоковых реакциях. У мышей вирусная экспрессия активированного TAZ способствует воспалению в печени, НАЖБП и образованию опухолей, связывая TAZ - но не YAP - с признаком воспаления в развитии опухоли (Wang et al., 2016; Hagenbeek et al., 2018). Механистически, TAZ / TEAD напрямую активирует передачу сигналов Hedgehog с повышенной экспрессией про-фибриногенных факторов в HSC, включая остеопонтин, Timp1 и Col1a1 (Machado et al.2015; Wang et al., 2016). Интересно, что скрининг транспозонов у мышей выявил мутации в адаптере белка Hippo Sav1 в опухолях, ассоциированных с НАЖБП, но не тех, которые развивались при вирусном гепатите (Kodama et al., 2018). Один из механизмов, с помощью которого стеатоз может способствовать активации YAP / TAZ, связан с белком JCAD, связанным с ожирением, который ингибирует LATS2 выше по течению (Ye et al., 2017). На сегодняшний день дифференциальная роль YAP и TAZ далека от понимания, и большинство моделей заболеваний печени сосредоточены на роли YAP.Тем не менее, вышеупомянутые исследования показывают, что TAZ может играть уникальную роль в стеатогепатите и развитии фиброза, связанного с НАЖБП. Цирроз и хроническое воспаление в печени являются ведущими факторами риска развития рака печени (Massarweh and El-Serag, 2017). Хотя активация YAP является ранним событием в развитии HCC (Perra et al., 2014), мутации в пути Hippo редки - что указывает на альтернативные регуляторные пути для стимулирования передачи сигналов YAP.Поиск восходящих регуляторов передачи сигналов Hippo привел к выявлению множества регуляторных механизмов, начиная от механотрансдукции по передаче сигналов GPCR до взаимодействующих путей, наиболее заметно пути передачи сигналов Wnt (Meng et al., 2016; Dobrokhotov et al., 2018; Liu et al. al., 2018). Учитывая увеличение жесткости печени, которое происходит при циррозе, механические воздействия являются ключевыми кандидатами для онкогенной активации YAP, которая способствует прогрессированию фиброза и раку печени.Механистически было показано, что некоторые мембранно-ассоциированные белки, такие как Nf2 / Merlin, строительные леса семейства ангиомотинов и белки WWC, положительно и - для некоторых членов семейства ангиомотинов - негативно регулируют передачу сигналов Hippo (Zhang et al., 2010; Li et al., 2012; Yi et al., 2013; Moleirinho et al., 2017; Hermann et al., 2018) - однако актуальность этих белков при циррозе печени до настоящего времени неизвестна. Несколько взаимодействующих путей, включая передачу сигналов Wnt и Notch, играют роль в контроле онкогенного YAP при раке печени.При гепатоцеллюлярной карциноме передача сигналов Notch не только активируется YAP, но также поддерживает петлю положительной обратной связи в Mst1 / 2-дефицитной печени для усиления передачи сигналов YAP и онкогенеза. Напротив, передача сигналов Wnt, как было показано, ингибирует передачу сигналов YAP / TAZ в печени - частично путем вмешательства в петлю обратной связи Notch-YAP (Tschaharganeh et al., 2013; Kim et al., 2017b, c). Интересно, что инактивирующие мутации в негативном регуляторе Wnt AXIN не приводят к усилению передачи сигналов Wnt / β-catenin в гепатоцеллюлярной карциноме, как можно было бы ожидать, а вместо этого связаны с пролиферативным фенотипом и сигнатурными генами, обогащенными для передачи сигналов Notch и YAP, указывающими на Wnt -независимое прямое или косвенное ингибирование YAP с помощью AXIN (Abitbol et al.2018). Вниз по течению от активированного YAP / TAZ, неконтролируемая пролиферация и нерегулируемый контроль клеточного цикла являются одним из ключевых механизмов, стимулирующих развитие опухоли. Несколько исследований показали, что YAP / TEAD взаимодействуют с факторами транскрипции E2F ниже передачи сигналов ретинобластомы, чтобы способствовать пролиферации при раке, в том числе при раке печени (Ehmer et al., 2014; Kapoor et al., 2014; Ehmer and Sage, 2015; Fitamant et al., 2015; Hiemer et al., 2015; Shi et al., 2016). Хотя низкие уровни активного YAP недостаточны для индукции пролиферации в спокойной печени, сопутствующая активация других пролиферативных сигналов при повреждении или воспалении печени дает YAP-активированным гепатоцитам пролиферативное преимущество, приводящее к их размножению (Ehmer et al.2014; Су и др., 2015). Помимо пролиферации, передача сигналов YAP в HCC участвует во множестве связанных с раком путей, включая подавление апоптоза, дерегулируемый ответ ER / unfolded protein и хромосомную нестабильность (CNI) (Rosenbluh et al., 2012; Wu et al., 2015; Weiler et al., 2017). Наконец, что не менее важно, передача сигналов Hippo является важным игроком в регуляции воспалительной микроокружающей среды. В печени с дефицитом Mst1 / 2 мишень YAP Mcp1 запускает накопление инфильтрирующих опухоль макрофагов, которые нарушают иммунный клиренс трансформированных гепатоцитов и способствуют развитию ГЦК (Guo et al., 2017; Kim et al., 2018). Если возрастающая роль передачи сигналов Hippo в противораковом иммунитете имеет какое-либо отношение к заболеванию печени, еще предстоит изучить, но она может сыграть важную роль в оптимизации стратегий иммунотерапии, направленной на ГЦК (Taha et al., 2018). Эффективное нацеливание на передачу сигналов Hippo in vivo - либо для стимуляции регенерации печени, либо для ингибирования фиброза и прогрессирования рака - представляет неудовлетворенную потребность в заболевании печени.Печеночная инактивация MST-киназ или YAP с помощью микроинкапсулированной siRNA , а также молчание YAP с помощью AAV-доставленной shRNA дали многообещающую эффективность на мышиных моделях, но переход в клинику пока не предвидится (Fitamant et al., 2015; Yin et al., 2016; Loforese et al., 2017; Jiang et al., 2018). Недавно транзиторная активация передачи сигналов YAP / TAZ с улучшенной регенерацией печени мыши была достигнута путем ингибирования MST1 / 2 киназ с использованием нового соединения XMU-MP-1 (Fan et al., 2016). В отличие от регенерации, ингибирование YAP / TAZ является перспективным терапевтическим подходом при циррозе, стеатогепатите или гепатоцеллюлярной карциноме. Результаты активации YAP в печени в основном зависят от ее взаимодействия с факторами транскрипции TEAD, что делает комплексы YAP / TEAD многообещающей мишенью. Вертепорфин, вещество, идентифицированное в скрининге соединений для ингибиторов YAP / TEAD, препятствовало развитию ГЦК в печени с дефицитом Nf2 (Liu-Chittenden et al., 2012).Тем не менее, клеточная токсичность из-за нецелевых эффектов, а также образование реактивных кислородных радикалов при световой активации препятствовали переходу вертепорфина в терапию рака (Konstantinou et al., 2017). Недавно антипаразитарный ивермектин был идентифицирован как ингибитор YAP / TEAD-зависимой транскрипции и был успешно использован для ингибирования активации YAP и избыточного роста печени при Mob1a / 1b-дефицитной печени. Хотя механизм ингибирования YAP ивермектином до настоящего времени не до конца понятен, соединение, по-видимому, имеет хороший профиль безопасности, что делает его перспективным лекарственным средством для ингибирования YAP in vivo (Nishio et al., 2016). В клетках аденокарциномы пищевода CA3 - новый ингибитор YAP / TEAD-зависимой транскрипции - успешно снижал рост опухолевых клеток вместе с установленной химиотерапией (Song et al., 2018). Однако это соединение до сих пор не было протестировано в печени. В дополнение к прямым ингибиторам YAP / TAZ, соединения, которые нацелены на восходящие регуляторы передачи сигналов Hippo, были в центре внимания нескольких исследований (Bae et al., 2017). В клетках HCC ингибиторы танкиразы, которые активируют членов семейства ангиомоксии, а также ингибиторы аурокиназы ограничивают рост опухолевых клеток путем модификации активности YAP (Jia et al., 2017; Лю и др., 2017). Хотя соединения, которые препятствуют механотрансдукции путем нацеливания на F-актин или Rho, успешно использовались для модификации передачи сигналов Hippo и активности YAP in vitro , их эффективность в клетках печени неизвестна, и токсичность, связанная с вмешательством цитоскелета, вероятно, ограничит их in vivo используют (Bae et al., 2017). В Wnt-активированных клетках рака толстой кишки нацеливание на YES1 с одобренным соединением дазатинибом ингибирует фосфорилирование тирозина YAP и активацию транскрипционного комплекса β-catenin-YAP1-TBX5 (Rosenbluh et al., 2012), в то время как в клетках молочной железы ингибирование PI3K / PDK1 способно сдерживать связывание PDK1 с SAV1, что приводит к подавлению передачи сигналов Hippo вверх по течению и снижению активности YAP (Fan et al., 2013). Опять же, эффективность этих соединений при раке печени не была исследована, и тканевая специфичность YAP-взаимодействующих путей сделает идентификацию направленных регуляторов выше по течению еще более сложной. В процессе разработки нескольких многообещающих препаратов-кандидатов для ингибирования YAP / TEAD их применение в настоящее время ограничено доклиническими исследованиями, и первые клинические испытания, которые ожидаются в ближайшем будущем, покажут, оправдаются ли огромные ожидания в отношении активности YAP в лечении. человеческих болезней. Наши знания о передаче сигналов Hippo в различных типах клеток печени расширились за последние годы (суммировано на рисунке 2), но важные вопросы остаются. При хроническом и остром повреждении печени мы до сих пор не полностью осознаем роль передачи сигналов Hippo во взаимодействии между HSC, LSEC, клетками желчных протоков и гепатоцитами. Помимо передачи сигналов ежу, существуют ли другие пути, такие как передача сигналов TGFbeta, и могут ли эти пути быть целевыми? С терапевтической модификацией передачи сигналов Hippo еще в зачаточном состоянии остается неясным, может ли ингибирование YAP или TAZ при стеатогепатите, циррозе и раке быть вредным для основного заболевания печени, поскольку оно может ингибировать регенерацию при повреждении клеток печени.Следовательно, было бы более разумно нацеливаться на специфические регуляторы воспаления или опухоли выше по течению передачи сигналов Hippo. Тем не менее, механизмы, которые регулируют печеночный путь гиппопотама во время развития печени, при гомеостазе печени во взрослой печени или при заболевании печени, не совсем понятны. Кроме того, клеточные различия в результатах активации YAP и TAZ еще предстоит изучить. В этом контексте особое значение имеет то, что роль YAP и TAZ не является полностью избыточной, и расшифровка отдельных функций обоих ортологов, несомненно, поможет понять роль передачи сигналов Hippo в печени.Для решения всех этих вопросов в сложной микроокружающей среде печени необходимы дальнейшие исследования. Рисунок 2. Сигнал гиппопотама в печени. ГЦК, гепатоцеллюлярная карцинома; ICC, внутрипеченочная холангиокарцинома. SM и UE составили и написали рукопись. Работа была профинансирована Вильгельмом Сандером-Стифтунгом, Германия (UE, грант №: 2017.115.1). Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов. Abitbol S., Dahmani R., Coulouarn C., Ragazzon B., Mlecnik B., Senni N. и др. (2018). Дефицит AXIN в гепатоцитах человека и мыши вызывает гепатоцеллюлярную карциному в отсутствие активации бета-катенина. J. Hepatol. 68, 1203–1213. doi: 10.1016 / j.jhep.2017.12.018 PubMed Аннотация | CrossRef Полный текст | Google Scholar Alder, О., Cullum, R., Lee, S., Kan, A.C., Wei, W., Yi, Y., et al. (2014). Передача сигналов Hippo влияет на переключение энхансеров HNF4A и FOXA2 во время дифференцировки гепатоцитов. Cell Rep. 9, 261–271. doi: 10.1016 / j.celrep.2014.08.046 PubMed Аннотация | CrossRef Полный текст | Google Scholar Anakk, S., Bhosale, M., Schmidt, V.A., Johnson, R.L., Finegold, M.J. and Moore, D.D. (2013). Желчные кислоты активируют YAP, способствуя канцерогенезу в печени. Cell Rep. 5, 1060–1069. doi: 10.1016 / j.celrep.2013.10.030 PubMed Аннотация | CrossRef Полный текст | Google Scholar Bai, H., Zhang, N., Xu, Y., Chen, Q.Хан М., Поттер Дж. Дж. И др. (2012). Связанный с Да белок регулирует печеночный ответ после перевязки желчных протоков. Гепатология 56, 1097–1107. doi: 10.1002 / hep.25769 PubMed Аннотация | CrossRef Полный текст | Google Scholar Benhamouche, S., Curto, M., Saotome, I., Gladden, A.B., Liu, C.H., Giovannini, M., et al. (2010). Nf2 / Merlin контролирует гомеостаз предшественников и онкогенез в печени. Genes Dev. 24, 1718–1730. doi: 10.1101 / gad.1938710 PubMed Аннотация | CrossRef Полный текст | Google Scholar Benhamouche-Trouillet, S., O’Loughlin, E., Liu, C.H., Polacheck, W., Fitamant, J., McKee, M., et al. (2018). Независимая от пролиферации роль NF2 (мерлина) в ограничении желчного морфогенеза. Разработка 145: dev162123. doi: 10.1242 / dev.162123 PubMed Аннотация | CrossRef Полный текст | Google Scholar Камарго, Ф. Д., Гохале, С., Джоннидис, Дж. Б., Фу, Д., Белл, Г. В., Джениш, Р., и другие. (2007). YAP1 увеличивает размер органа и расширяет недифференцированные клетки-предшественники. Curr. Biol. 17, 2054–2060. doi: 10.1016 / j.cub.2007.10.039 PubMed Аннотация | CrossRef Полный текст | Google Scholar Dechene, A., Sowa, J.P., Gieseler, R.K., Jochum, C., Bechmann, L.P., El Fouly, A., et al. (2010). Острая печеночная недостаточность связана с повышенной жесткостью печени и активацией звездчатых клеток печени. Гепатология 52, 1008–1016. дои: 10,1002 / геп.23754 PubMed Аннотация | CrossRef Полный текст | Google Scholar Ding, B.S., Cao, Z., Lis, R., Nolan, D.J., Guo, P., Simons, M., et al. (2014). Дивергентные ангиокринные сигналы от сосудистой ниши уравновешивают регенерацию печени и фиброз. Природа 505, 97–102. doi: 10.1038 / nature12681 PubMed Аннотация | CrossRef Полный текст | Google Scholar Доброхотов О., Самсонов М., Сокабе М. и Хирата Х. (2018). Механорегуляция и патология YAP / TAZ через механизмы Hippo и non-Hippo. Clin. Перев. Med. 7:23. doi: 10.1186 / s40169-018-0202-9 PubMed Аннотация | CrossRef Полный текст | Google Scholar Донг Дж., Фельдманн Г., Хуан Дж., Ву С., Чжан Н., Комерфорд С.А. и др. (2007). Выяснение универсального механизма контроля размера у дрозофилы и млекопитающих. Cell 130, 1120–1133. doi: 10.1016 / j.cell.2007.07.019 PubMed Аннотация | CrossRef Полный текст | Google Scholar Дюпон С., Морсут Л., Арагона М.Enzo E., Giulitti S., Cordenonsi M. et al. (2011). Роль YAP / TAZ в механотрансдукции. Природа 474, 179–183. doi: 10.1038 / nature10137 PubMed Аннотация | CrossRef Полный текст | Google Scholar Ehmer, U., Zmoos, A.F., Auerbach, R.K., Vaka, D., Butte, A.J., Kay, M.A., et al. (2014). Контроль размера органа доминирует над инактивацией семейства Rb для ограничения пролиферации in vivo. Cell Rep. 8, 371–381. doi: 10.1016 / j.celrep.2014.06.025 PubMed Аннотация | CrossRef Полный текст | Google Scholar Fan, F.He, Z., Kong, L.L., Chen, Q., Yuan, Q., Zhang S., et al. (2016). Фармакологическое воздействие на киназы MST1 и MST2 способствует восстановлению и регенерации тканей. Sci. Перев. Med. 8: 352ра108. doi: 10.1126 / scitranslmed.aaf2304 PubMed Аннотация | CrossRef Полный текст | Google Scholar Fan, R., Kim, N.G. и Gumbiner, B.M. (2013). Регуляция пути Бегемота митогенными факторами роста с помощью фосфоинозитид-3-киназы и фосфоинозитид-зависимой киназы-1. Proc.Natl. Акад. Sci. США 110, 2569-2574. doi: 10.1073 / pnas.1216462110 PubMed Аннотация | CrossRef Полный текст | Google Scholar Fitamant, J., Kottakis, F., Benhamouche S., Tian, H. S., Chuvin, N., Parachoniak, C.A., et al. (2015). Ингибирование YAP восстанавливает дифференцировку гепатоцитов при прогрессирующем ГЦК, приводя к регрессии опухоли. Cell Rep. doi: 10.1016 / j.celrep.2015.02.027 [Epub перед печатью]. PubMed Аннотация | CrossRef Полный текст | Google Scholar Font-Burgada, J., Shalapour S., Ramaswamy S., Hsueh B., Rossell D., Umemura A., et al. (2015). Гибридные перипортальные гепатоциты регенерируют поврежденную печень, не вызывая рака. Cell 162, 766–779. doi: 10.1016 / j.cell.2015.07.026 PubMed Аннотация | CrossRef Полный текст | Google Scholar
4.7 Размер и заряд
4.8 Базовый характер
4.9 Концентрация лиганда
4.10 Хелатирующий эффект
Рисунок 1.
Рисунок 2.
Рисунок 3.
4.11 Размер кольца
4.12 Количество колец
Ион металла log βMY (25 ° C, I = 0,1 М) Ca 2+ 11 ,2 Cu 2+ 19,8 Fe 3+ 24,9 Таблица 1.
4.13 Стерический эффект
4.14 Макроциклический эффект
Рисунок 4.
Почему печень такая удивительная? · Границы для молодых умов
Аннотация
Строение и функции печени
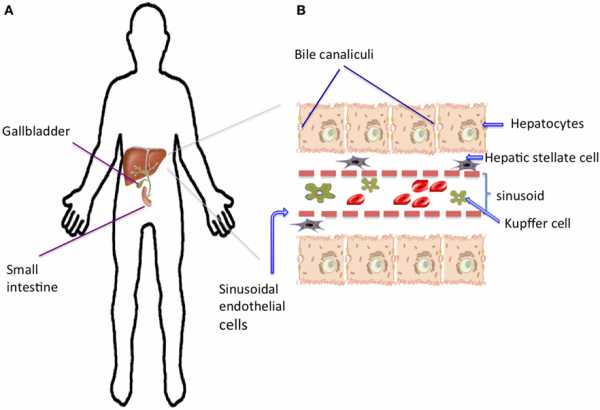
Что такое регенерация печени?
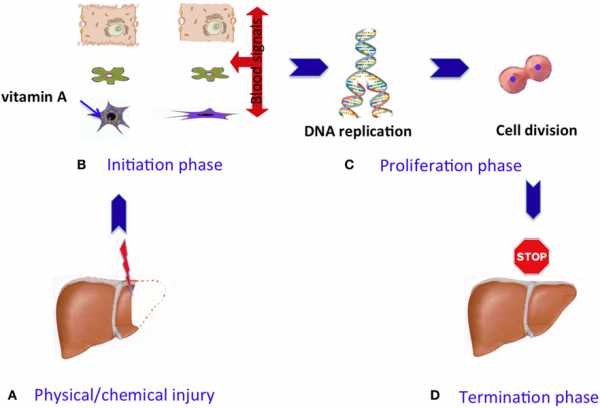
Почему полезно знание о регенерации печени?
Заключение
Глоссарий
Заявление о конфликте интересов
Благодарности
Список литературы
границ | Сигнализация гиппопотама в печени - длинная и вечная история
Сигнал бегемота млекопитающих - все началось в печени
регуляторов гиппопотама ограничивают пролиферацию и поддерживают дифференцировку в гепатоцитах
Сигналы гиппопотама в клетках желчных протоков - одинаковые, но разные
Регенерация печени - на хорошей стороне YAP / TAZ
Фиброз печени - рубцовое лицо YAP и TAZ
Стеатогепатит - связь с метаболизмом и воспалением
Рак печени - онкогенные корни YAP
Таргетирование сигналов гиппопотама при заболевании печени
перспективы на будущее
Вклад автора
Финансирование
Заявление о конфликте интересов
Отзывы