





2011-2017 © МБУЗ ГКП № 7, г.Челябинск.
Обмен билирубина
В основе образования билирубина лежит разрушение железосодержащей части гемоглобина и других гемсодержащих белков и ферментов. Гем распадается до биливердина, который восстанавливается в билирубин.
Свободный билирубин токсичен, не растворяются в воде и циркулирует в крови в комплексе с альбуминами. Этот билирубин дает непрямую реакцию Ван ден Берга (после осаждения альбуминов спиртом), поэтому называется непрямым.
Непрямой билирубин, будучи связанным с альбуминами, не проходит через неповрежденные мембраны почечных клубочков и не фильтруется в мочу.
Выведение билирубина осуществляется с желчью через кишечник. Билирубин, связанный с альбуминами, доставляется кровью в печень. Билирубин легко проникает через мембраны гепатоцитов, альбумины остаются в кровотоке.
В гепатоцитах билирубин соединяется с глюкуроновой кислотой, превращаясь в билирубинмоно- и диглюкуронид («выпрямляется», прямой,связанный).
Образованные билирубинглюкурониды нетоксичны, легко растворимы. Они направляются с желчью в кишечник для выведения из организма.
Из кишечника билирубин глюкурониды частично поступают в кровоток и, находясь в крови, представляют собой фракцию прямого билирубина, который дает прямую реакцию Ван ден Берга. Прямой билирубин в отличие от непрямого легко проникает через почечные фильтры и может выделяться с мочой.
В физиологических условиях сыворотка крови содержит при мерно 25 % прямого билирубина (связанного с глюкуроновой кислотой) и 75 % непрямого билирубина (альбумин-билирубина).
Таким образом, общий билирубин крови представляет собой суммарное количество непрямого и прямого билирубина.
У здоровых людей в сыворотке крови содержится билирубина 1,7—20,5 мкмоль/литр; прямого — 0,4—5,1 мкмоль/литр.
Обмен билирубина
Билирубин представляет собой конечный продукт распада гема.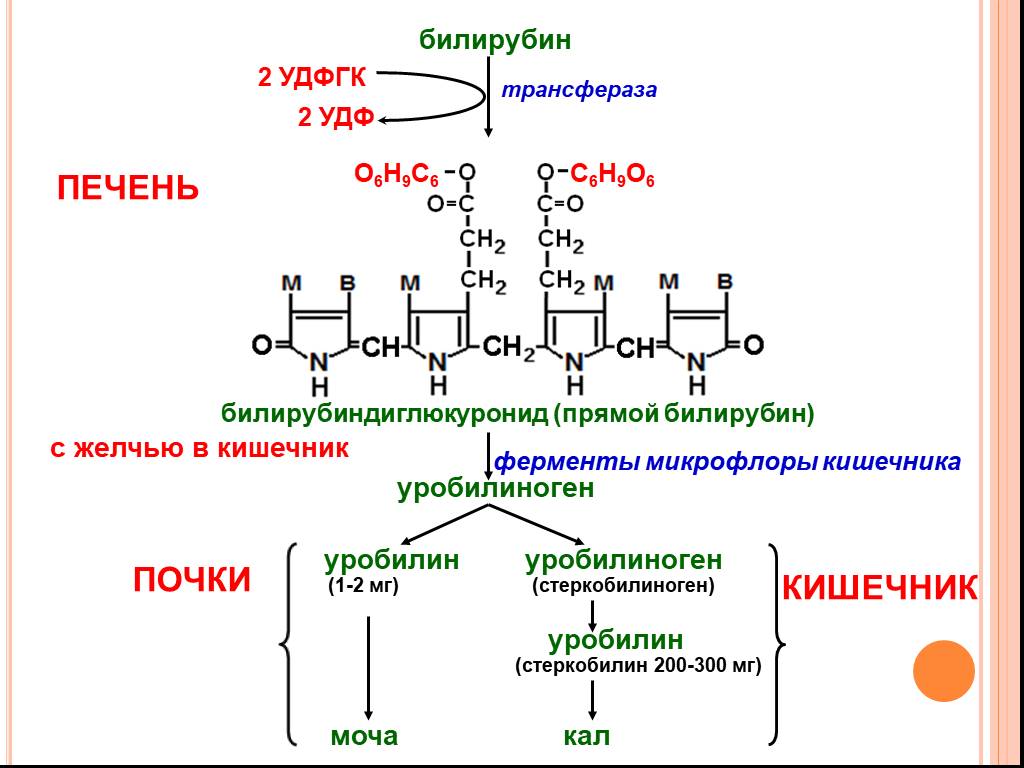 Основная часть (80—85%) билирубина образуется из гемоглобина и лишь небольшая часть — из других гемсодержащих белков, например цитохрома Р450. Образование билирубина происходит в клетках ретикулоэндотелиальной системы. Ежедневно образуется около 300 мг билирубина.
Основная часть (80—85%) билирубина образуется из гемоглобина и лишь небольшая часть — из других гемсодержащих белков, например цитохрома Р450. Образование билирубина происходит в клетках ретикулоэндотелиальной системы. Ежедневно образуется около 300 мг билирубина.
Преобразование гема в билирубин происходит с участием микросомального фермента гемоксигеназы, для работы которого требуются кислород и НАДФН. Расщепление порфиринового кольца происходит селективно в области метановой группы в положении а. Атом углерода, входящий в состав a-метанового мостика, окисляется до моноксида углерода, и вместо мостика образуются 2 двойные связи с молекулами кислорода, поступающими извне. Образующийся в результате этого линейный тетрапиррол по структуре является IX-aльфа-биливердином. Далее он преобразуется биливердинредуктазой, цитозольным ферментом, в IX-aльфа-билирубин. Линейный тетрапиррол такой структуры должен растворяться в воде, в то время как билирубин является жирорастворимым веществом. Растворимость в липидах определяется структурой IX-aльфа-билирубина — наличием 6 стабильных внутримолекулярных водородных связей [5]. Эти связи можно разрушить спиртом в диазореакции (Ван ден Берга), в которой неконъюгированный (непрямой) билирубин превращается в конъюгированный (прямой). In vivo стабильные водородные связи разрушаются этерификацией с помощью глюкуроновой кислоты.
Растворимость в липидах определяется структурой IX-aльфа-билирубина — наличием 6 стабильных внутримолекулярных водородных связей [5]. Эти связи можно разрушить спиртом в диазореакции (Ван ден Берга), в которой неконъюгированный (непрямой) билирубин превращается в конъюгированный (прямой). In vivo стабильные водородные связи разрушаются этерификацией с помощью глюкуроновой кислоты.
Около 20% циркулирующего билирубина образуется не из гема зрелых эритроцитов, а из других источников. Небольшое количество поступает из незрелых клеток селезёнки и костного мозга. При гемолизе это количество увеличивается. Остальной билирубин образуется в печени из гемсодержащих белков, например миоглобина, цитохромов, и из других неустановленных источников. Эта фракция увеличивается при пернициозной анемии, эритропоэтической уропорфирин и при синдроме Криглера-Найяра.
Транспорт и конъюгация билирубина в печени
Неконъюгированный билирубин в плазме прочно связан с альбумином.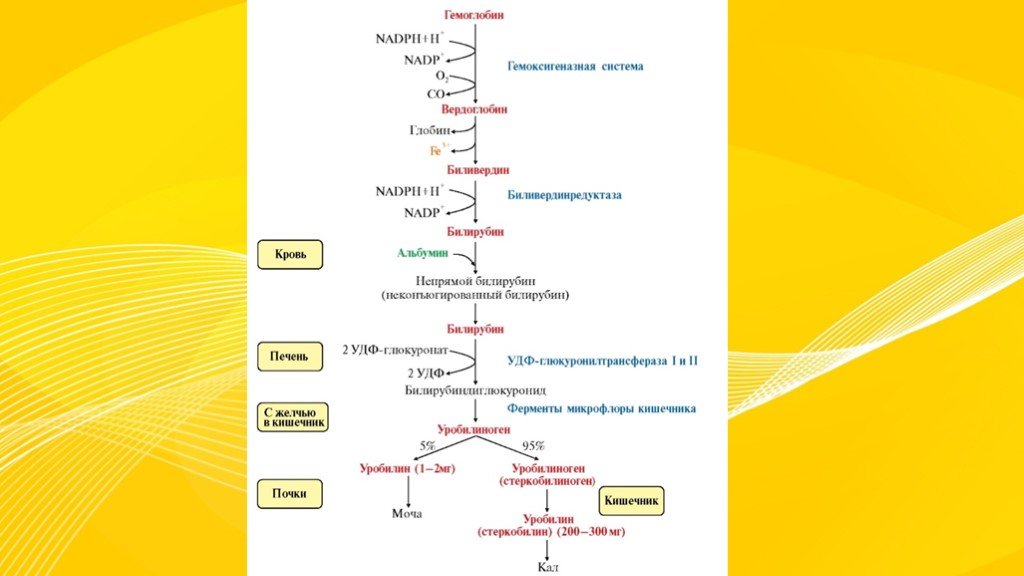 Только очень небольшая часть билирубина способна подвергаться диализу, однако под влиянием веществ, конкурирующих с билирубином за связывание с альбумином (например, жирных кислот или органических анионов), она может увеличиваться. Это имеет важное значение у новорождённых, у которых ряд лекарств (например, сульфаниламиды и салицилаты) может облегчать диффузию билирубина в головной мозг и таким образом способствовать развитию ядерной желтухи.
Только очень небольшая часть билирубина способна подвергаться диализу, однако под влиянием веществ, конкурирующих с билирубином за связывание с альбумином (например, жирных кислот или органических анионов), она может увеличиваться. Это имеет важное значение у новорождённых, у которых ряд лекарств (например, сульфаниламиды и салицилаты) может облегчать диффузию билирубина в головной мозг и таким образом способствовать развитию ядерной желтухи.
Печенью выделяются многие органические анионы, в том числе жирные кислоты, жёлчные кислоты и другие компоненты жёлчи, не относящиеся к жёлчным кислотам, такие как билирубин (несмотря на его прочную связь с альбумином). Исследования показали, что билирубин отделяется от альбумина в синусоидах, диффундирует через слой воды на поверхности гепатоцита |55]. Высказанные ранее предположения о наличии рецепторов альбумина не подтвердились. Перенос билирубина через плазматическую мембрану внутрь гепатоцита осуществляется с помощью транспортных белков, например транспортного белка органических анионов [50], и/или по механизму «флип-флоп» [55].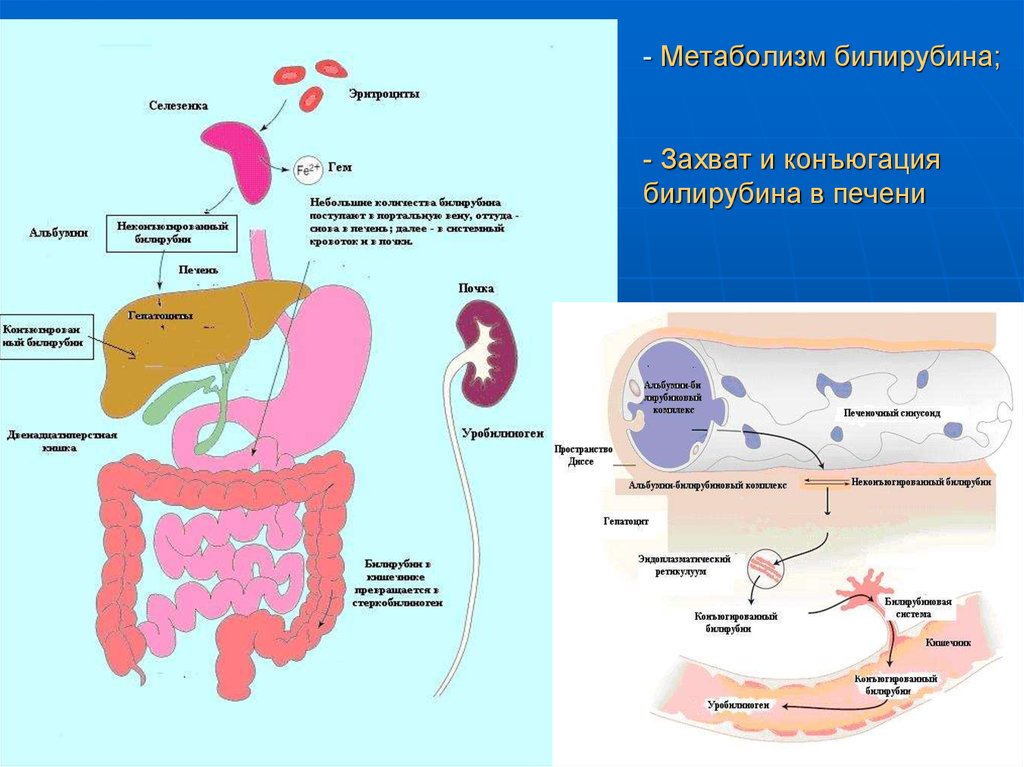 Захват билирубина высокоэффективен благодаря его быстрому метаболизму в печени в реакции глюкуронидизации и выделению в жёлчь, а также вследствие наличия в цитозоле связывающих белков, таких как лигандины (глутатион-8-трансфераза).
Захват билирубина высокоэффективен благодаря его быстрому метаболизму в печени в реакции глюкуронидизации и выделению в жёлчь, а также вследствие наличия в цитозоле связывающих белков, таких как лигандины (глутатион-8-трансфераза).
Неконъюгированный билирубин представляет собой неполярное (жирорастворимое) вещество. В реакции конъюгации он превращается в полярное (водорастворимое вещество) и может благодаря этому выделяться в желчь. Эта реакция протекает с помощью микросомального фермента уридиндифосфатглюкуронилтрансферазы (УДФГТ), превращающего неконъюгированный билирубин в конъюгированный моно- и диглюкуронид билирубина. УДФГТ является одной из нескольких изоформ фермента, обеспечивающих конъюгацию эндогенных метаболитов, гормонов и нейротрансмиттеров.
Ген УДФГТ билирубина находится на 2-й паре хромосом. Структура гена сложная (рис. 12-4) [2, 54]. У всех изоформ УДФГТ постоянными компонентами являются экзоны 2—5 на 3'-конце ДНК гена. Для экспрессии гена необходимо вовлечение одного из нескольких первых экзонов.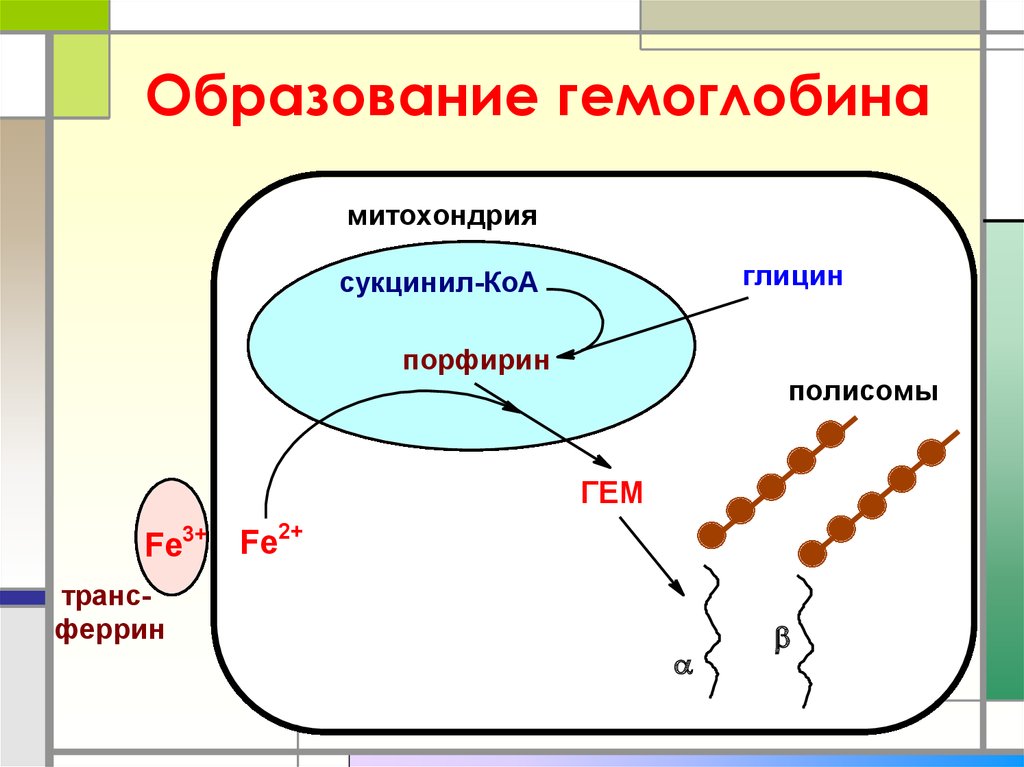 Так, для образования изоферментов билирубин-УДФГТ1*1 и 1*2 необходимо вовлечение соответственно экзонов 1А и ID. Изофермент 1*1 участвует в конъюгации практически всего билирубина, а изофермент 1*2 почти или вовсе не участвует в этом [25]. Другие экзоны (IF и 1G) кодируют изоформы фенол-УДФГТ. Таким образом, выбор одной из последовательностей экзона 1 определяет субстратную специфичность и свойства ферментов.
Так, для образования изоферментов билирубин-УДФГТ1*1 и 1*2 необходимо вовлечение соответственно экзонов 1А и ID. Изофермент 1*1 участвует в конъюгации практически всего билирубина, а изофермент 1*2 почти или вовсе не участвует в этом [25]. Другие экзоны (IF и 1G) кодируют изоформы фенол-УДФГТ. Таким образом, выбор одной из последовательностей экзона 1 определяет субстратную специфичность и свойства ферментов.
Дальнейшая экспрессия УДФГТ 1*1 зависит также от промоторного участка на 5'-конце, связанного с каждым из первых экзонов |6|. Промоторный участок содержит последовательность ТАТАА.
Детали строения гена важны для понимания патогенеза неконъюгированной гипербилирубинемии (синдромы Жильбера и Криглера—Найяра; см. соответствующие разделы), когда в печени содержание ферментов, ответственных за конъюгацию, снижено или они отсутствуют.
Активность УДФГТ при печёночно-клеточной желтухе поддерживается на достаточном уровне, а при холестазе даже увеличивается.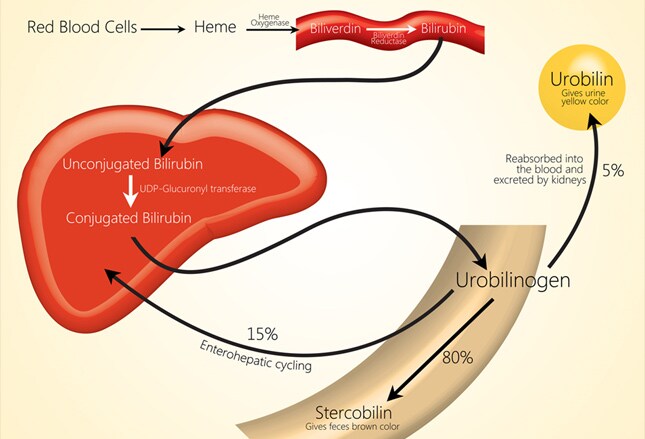 У новорождённых активность УДФГТ низкая.
У новорождённых активность УДФГТ низкая.
У человека в жёлчи билирубин представлен в основном д и глюкуронидом. Превращение билирубина в моноглюкуронид, а также в диглюкуронид происходит в одной и той же микросомальной системе глюкуронилтрансферазы [37]. При перегрузке билирубином, например при гемолизе, образуется преимущественно моноглюкуронид, а при уменьшении поступления билирубина или при индукции фермента возрастает содержание диглюкуронида.
Наиболее важное значение имеет конъюгация с глюкуроновой кислотой, однако небольшое количество билирубина конъюгируется с сульфатами, ксилозой и глюкозой; при холестазе эти процессы усиливаются [II].
В поздних стадиях холестатической или печёночно-клеточной желтухи, несмотря на высокое содержание в плазме, билирубин в моче не выявляется. Очевидно, причиной этого является образование билирубина типа III, моноконъюгированного, который ковалентно связан с альбумином [54]. Он не фильтруется в клубочках и, следовательно, не появляется в моче.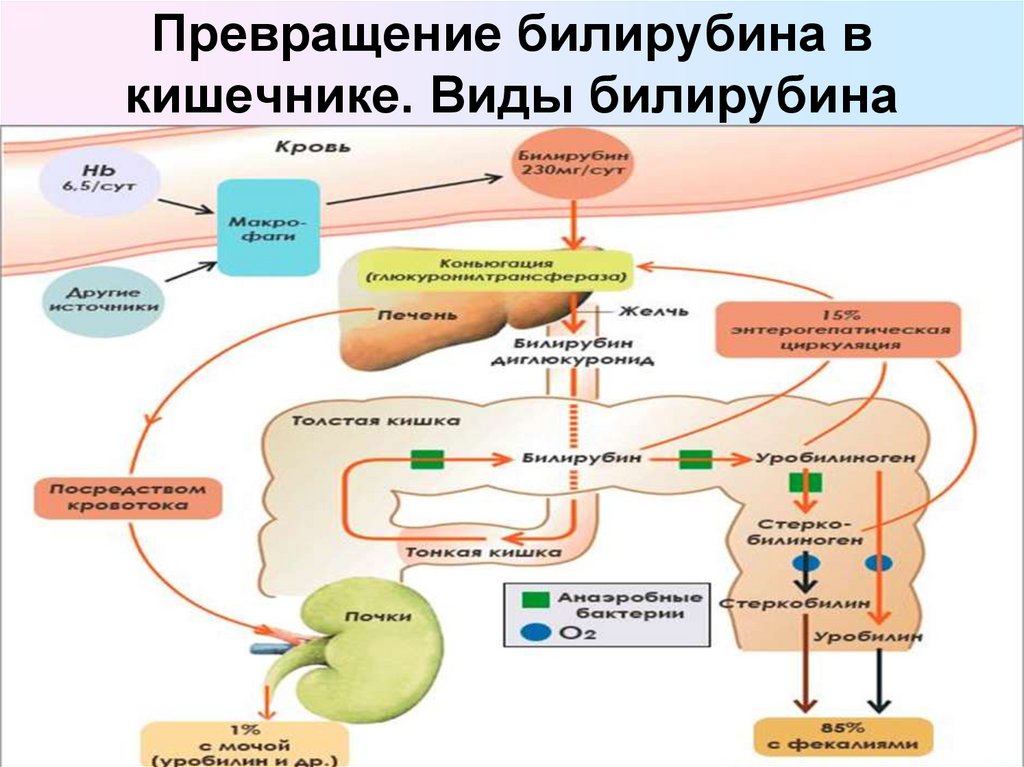 Это снижает практическую значимость проб, применяемых для определения содержания билирубина в моче.
Это снижает практическую значимость проб, применяемых для определения содержания билирубина в моче.
Экскреция билирубина в канальцы происходит с помощью семейства АТФ-зависимых мультиспецифичных транспортных белков для органических анионов [27]. Скорость транспорта билирубина из плазмы в жёлчь определяется этапом экскреции глюкуронида билирубина.
Жёлчные кислоты переносятся в жёлчь с помощью другого транспортного белка. Наличие разных механизмов транспорта билирубина и жёлчных кислот можно проиллюстрировать на примере синдрома Дубина—Джонсона, при котором нарушается экскреция конъюгированного билирубина, но сохраняется нормальная экскреция жёлчных кислот. Большая часть конъюгированного билирубина в жёлчи находится в смешанных мицеллах, содержащих холестерин, фосфолипиды и жёлчные кислоты. Значение аппарата Гольджи и микрофиламентов цитоскелета гепатоцитов для внутриклеточного транспорта конъюгированного билирубина пока не установлено.
Диглюкуронид билирубина, находящийся в жёлчи, водорастворим (полярная молекула), поэтому в тонкой кишке не всасывается.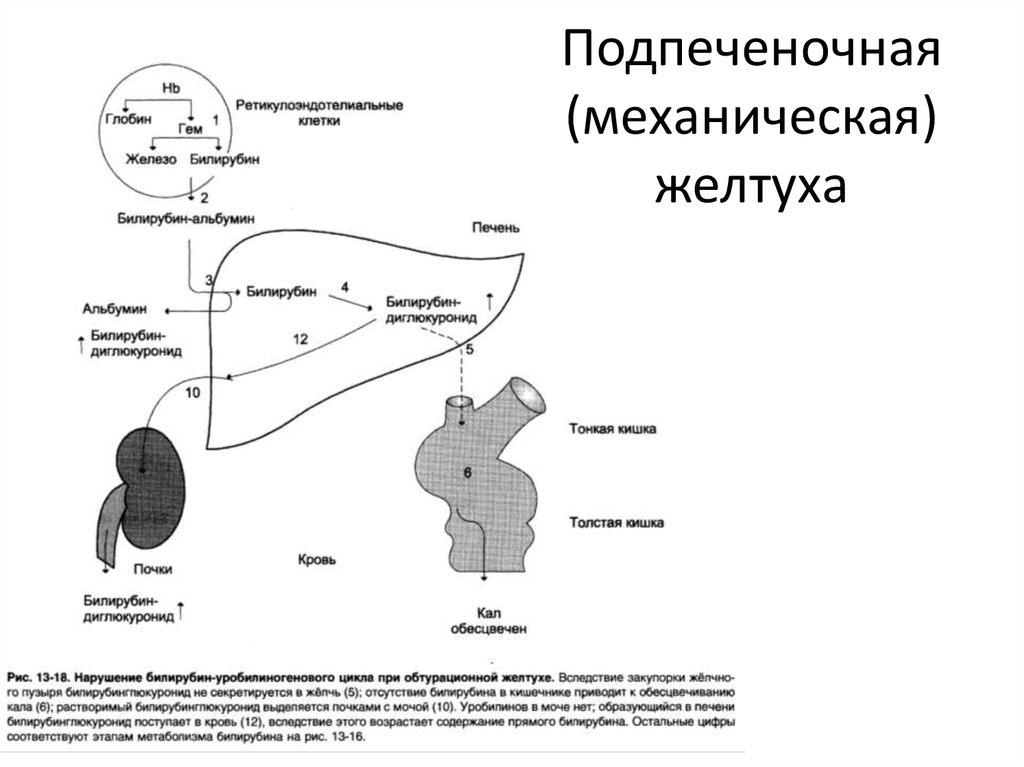 В толстой кишке конъюгированный билирубин подвергается гидролизу b-глюкуронидазами бактерий с образованием уробилиногенов. При бактериальном холангите часть диглюкуронида билирубина гидролизуется уже в жёлчных путях с последующей преципитацией билирубина. Этот процесс может иметь важное значение для образования билирубиновых жёлчных камней.
В толстой кишке конъюгированный билирубин подвергается гидролизу b-глюкуронидазами бактерий с образованием уробилиногенов. При бактериальном холангите часть диглюкуронида билирубина гидролизуется уже в жёлчных путях с последующей преципитацией билирубина. Этот процесс может иметь важное значение для образования билирубиновых жёлчных камней.
Уробилиноген, имея неполярную молекулу, хорошо всасывается в тонкой кишке и в минимальном количестве — в толстой. Небольшое количество уробилиногена, которое в норме всасывается, вновь экскретируется печенью и почками {энтерогепатическая циркуляция). При нарушении функции гепатоцитов печёночная реэкскреция уробилиногена нарушается и увеличивается почечная экскреция. Данный механизм объясняет уробилиногенурию при алкогольной болезни печени, при лихорадке, сердечной недостаточности, а также на ранних стадиях вирусного гепатита.
Распределение билирубина в тканях при желтухе
Циркулирующий билирубин, связанный с белком, с трудом проникает в тканевые жидкости с низким содержанием белка. Если количество белка в них увеличивается, желтуха становится более выраженной. Поэтому экссудаты обычно более желтушны, чем транссудаты.
Если количество белка в них увеличивается, желтуха становится более выраженной. Поэтому экссудаты обычно более желтушны, чем транссудаты.
Ксантохромия цереброспинальной жидкости более вероятна при менингите; классическим примером этому может служить болезнь Вейля (желтушный лептоспироз) с сочетанием желтухи и менингита.
У новорождённых может наблюдаться желтушное прокрашивание базальных ганглиев головного мозга (ядерная желтуха), обусловленное высоким уровнем неконъюгированного билирубина в крови, имеющего сродство к нервной ткани.
При желтухе содержание билирубинам цереброспинальной жидкости небольшое: одна десятая или одна сотая от уровня билирубина в сыворотке.
При выраженной желтухе внутриглазная жидкость может окрашиваться в жёлтый цвет, чем объясняется чрезвычайно редкий симптом — ксантопсия (больные видят окружающие предметы в жёлтом цвете).
При выраженной желтухе жёлчный пигмент появляется в моче, поте, семенной жидкости, молоке.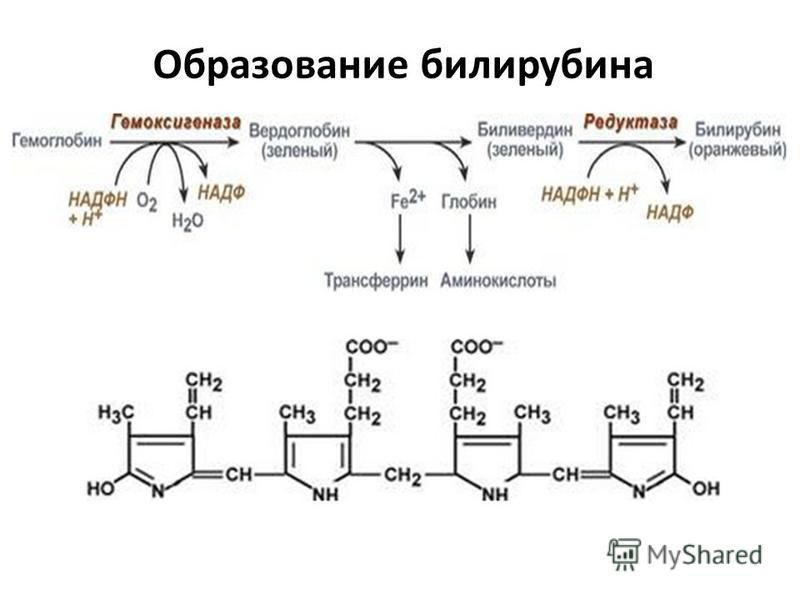 Билирубин является нормальным компонентом синовиальной жидкости, может содержаться и в норме.
Билирубин является нормальным компонентом синовиальной жидкости, может содержаться и в норме.
Цвет кожи парализованных и отёчных участков тела обычно не изменяется.
Билирубин легко связывается с эластической тканью. Она в большом количестве содержится в коже, склерах, стенке кровеносных сосудов, поэтому эти образования легко становятся желтушными. Этим же объясняется несоответствие выраженности желтухи и уровня билирубина в сыворотке в периоде выздоровления при гепатите и холестазе.
Факторы, определяющие выраженность желтухи
Даже при полной обструкции жёлчных путей выраженность желтухи может варьировать. Вслед за быстрым повышением уровень билирубина в сыворотке приблизительно через 3 нед начинает снижаться, даже если обструкция сохраняется. Выраженность желтухи зависит как от выработки жёлчного пигмента, так и от экскреторной функции почек. Скорость образования билирубина из гема может меняться; при этом возможно образование, помимо билирубина, и других продуктов, которые не вступают в диазореакцию.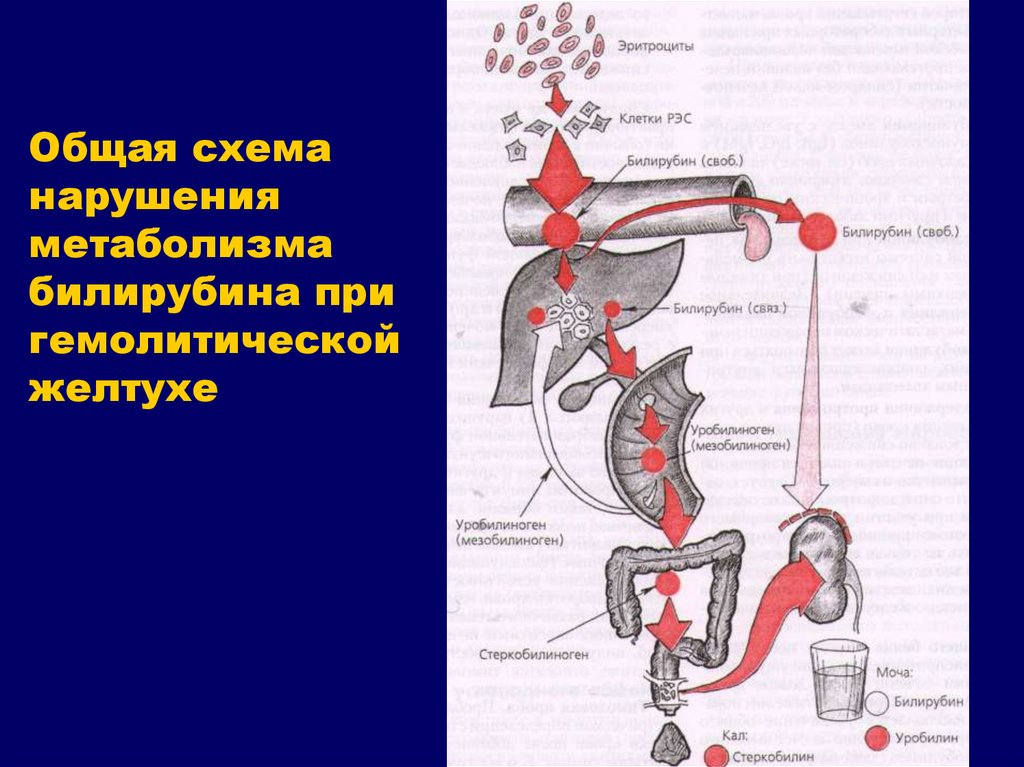 Билирубин, в основном неконъюгированный, может также выделяться из сыворотки слизистой оболочки кишечника.
Билирубин, в основном неконъюгированный, может также выделяться из сыворотки слизистой оболочки кишечника.
При длительном холестазе кожа приобретает зеленоватый оттенок, вероятно вследствие отложения биливердина, не участвующего в диазореакции (Ван ден Берга), а возможно, и других пигментов.
Конъюгированный билирубин, способный растворяться в воде и проникать в жидкости тела, вызывает более выраженную желтуху, чем неконъюгированный. Внесосудистое пространство тела больше, чем внутрисосудистое. Поэтому печёночно-клеточная и холестатическая желтуха обычно более интенсивная, чем гемолитическая.
Классификация желтухи
Существует 4 механизма развития желтухи.
Во-первых, возможно повышение нагрузки билирубином на гепатоциты. Во-вторых, могут нарушаться захват и перенос билирубина в гепатоцит. В-третьих, может нарушаться процесс конъюгации. И наконец, может нарушаться экскреция билирубина в жёлчь через канальцевую мембрану либо развиваться обструкция более крупных жёлчных путей.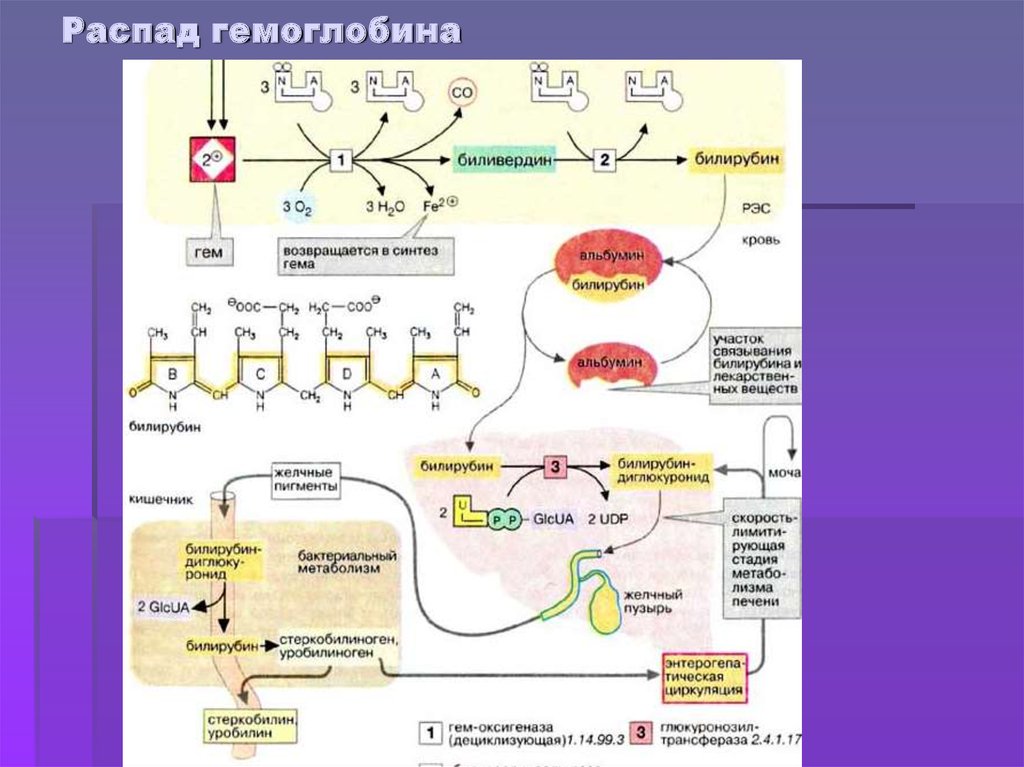
Выделяют 3 типа желтухи:
Эти типы желтухи, особенно печёночная и холестатическая, имеют во многом сходные проявления.
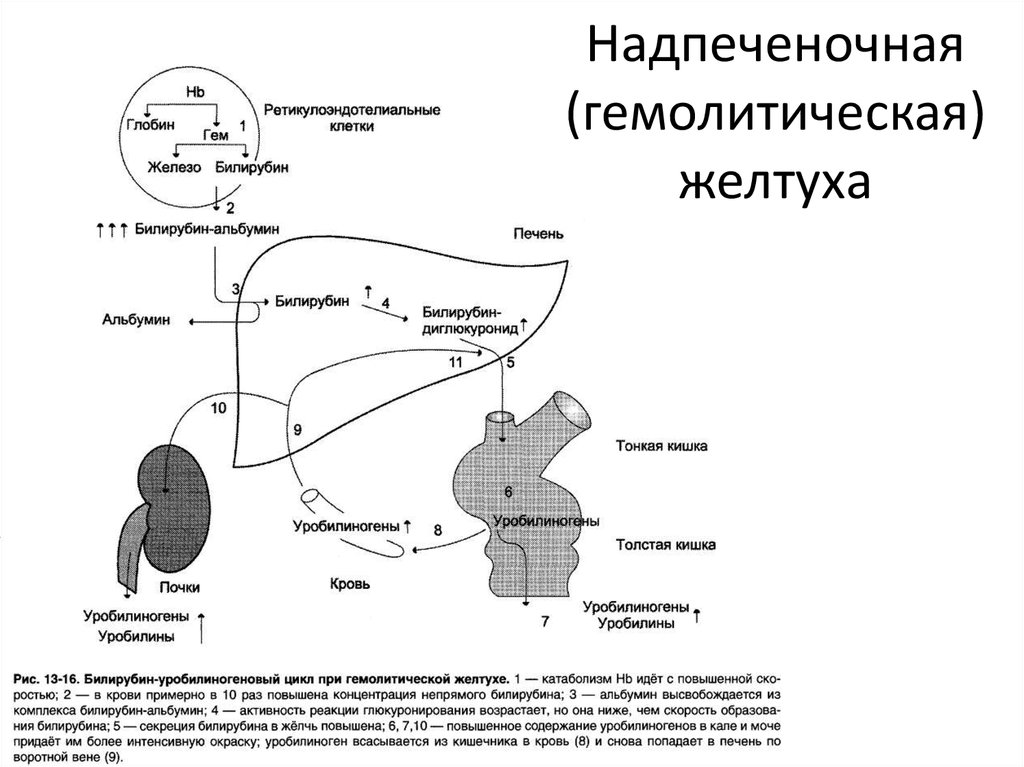
Надпечёночная желтуха. Уровень общего билирубина в сыворотке повышается, активность сывороточных трансаминаз и ЩФ сохраняется в пределах нормы. Билирубин представлен в основном неконъюгированной фракцией. В моче билирубин не выявляется. Этот тип желтухи развивается при гемолизе и наследственных нарушениях обмена билирубина.
Печёночная (печёночно-клеточная) желтуха (см. главы 16 и 18) обычно развивается быстро и имеет оранжевый оттенок. Больных беспокоят выраженная слабость и утомляемость. Печёночная недостаточность может быть выражена в разной степени. При лёгкой печёночной недостаточности можно выявить лишь незначительные нарушения психического статуса, более выраженная печёночная недостаточность сопровождается появлением «хлопающего» тремора, спутанности сознания и комы. Небольшая задержка жидкости может проявиться лишь увеличением массы тела, при значительной задержке жидкости появляются отёки и асцит. Вследствие нарушения синтеза печенью факторов свёртывания крови возможны кровоподтёки, как после венопункций, так и спонтанные. При биохимическом исследовании выявляют повышение активности сывороточных трансаминаз; при длительном течении заболевания возможно также снижение уровня альбумина в сыворотке.
Вследствие нарушения синтеза печенью факторов свёртывания крови возможны кровоподтёки, как после венопункций, так и спонтанные. При биохимическом исследовании выявляют повышение активности сывороточных трансаминаз; при длительном течении заболевания возможно также снижение уровня альбумина в сыворотке.
Холестатическая желтуха (см. главу 13) развивается при нарушении поступления жёлчи в двенадцатиперстную кишку. Значительного нарушения состояния больного (помимо симптомов основного заболевания) не происходит, отмечается интенсивный зуд. Желтуха прогрессирует, в сыворотке повышаются уровень конъюгированного билирубина, активность печёночной фракции ЩФ, ГГТП, а также уровень общего холестерина и конъюгированных жёлчных кислот. Вследствие стеатореи уменьшается масса тела и нарушается всасывание витаминов А, Д, Е, К, а также кальция.
Диагностика желтухи
Большое значение в установлении диагноза при желтухе имеют тщательно собранный анамнез, клиническое и лабораторное обследование и биохимический и клинический анализ крови.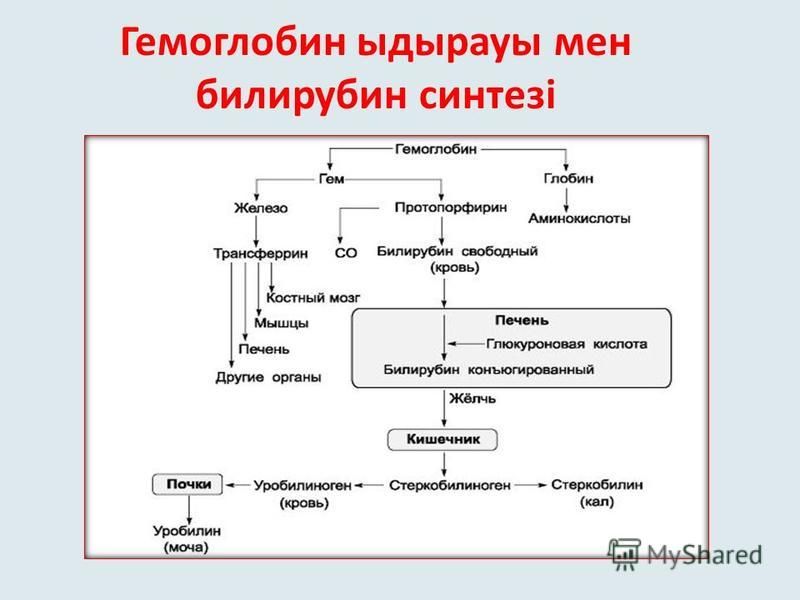 Необходимо исследование кала, которое должно включать анализ на скрытую кровь. При исследовании мочи следует исключить повышение содержания билирубина и уробилиногена. Дополнительные методы исследования — ультразвуковое исследование (УЗИ), биопсию печени и холангиографию (эндоскопическую или чрескожную) — применяют по показаниям в зависимости от типа желтухи.
Необходимо исследование кала, которое должно включать анализ на скрытую кровь. При исследовании мочи следует исключить повышение содержания билирубина и уробилиногена. Дополнительные методы исследования — ультразвуковое исследование (УЗИ), биопсию печени и холангиографию (эндоскопическую или чрескожную) — применяют по показаниям в зависимости от типа желтухи.
6
Синтез и выделение билирубина. Каждый эритроцит содержит примерно 270 миллионов молекул гемоглобина. Время жизни одного эритроцита 120 дней. 100-200 миллионов эритроцитов разрушаются каждый час. Процесс разрушения эритроцитов происходит в селезенке. В день разрушается 6 грамм гемоглобина.
Далее в макрофагах селезёнки происходит разрушение эритроцитов. Освобождается гемоглобин, который распадается на Гем и глобин. Гем в свою очередь распадается на двухвалентное железо, и неконъюгированный билирубин.
Гем выделяется из старых эритроцитов. В меньшей части из миоглобина, цитохромов, клеток костного мозга.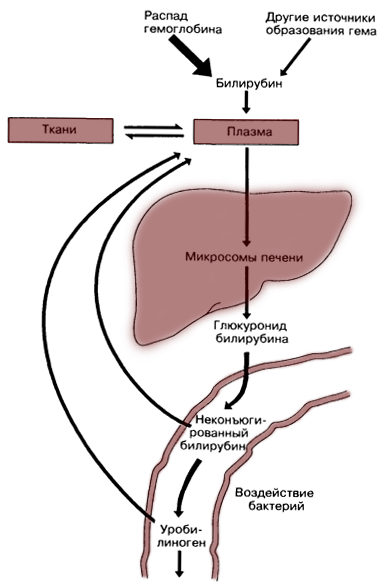 Макрофаги поглощают остаточный гем. С помощью гем оксидазы гем превращается в биливердин, а под воздействием биливердин редуктазы в билирубин.
Макрофаги поглощают остаточный гем. С помощью гем оксидазы гем превращается в биливердин, а под воздействием биливердин редуктазы в билирубин.
Структура Гема и Билирубина сходна, за исключением содержания двухвалентоного железа и разрыва одной связи.
Образовавшийся билирубин Токсичен. А за счёт того, что плохо растворим в воде, не может быть выведен из организма в таком виде. Переносится альбумином в печень.
Благодаря связыванию с альбумином неконъюгированный билирубин поступает в печень. В печени с помощью УДФ-глюкуронилтрансферазы происходит связывание неконъюгированного билирубина с глюкуроновой кислотой, образуется так называемый конъюгированный билирубин.
Глюкуроновая кислота имеет хорошую растворимость в воде, что делает билирубин хорошо растворимым в воде.
Если неконъюгированный билирубин связывается с одной молекулой глюкуроновой кислоты – образуется конъюгированный билирубин (моноглюкуронид), если с двумя — конъюгированный билирубин (диглюкуронид).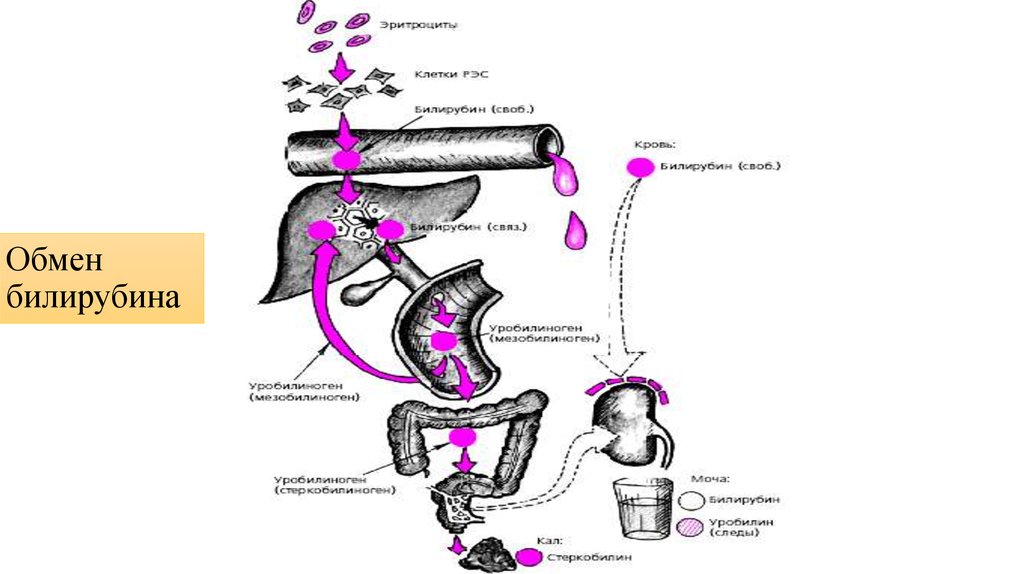
Конъюгация билирубина:
Происходит повышение водорастворимых свойств билирубина для экскреции с желчью.
Конъюгированный билирубин содержится в желчи, которая накапливается в жёлчном пузыре. Во время приёма пищи желчь поступает в тонкий кишечник.
Далее в толстом кишечнике начинается следующий этап преобразования билирубина. Под воздействием ферментов бактерий бета-глюкуронидаз происходит образование уробилиногена и отделение глюкуроновой кислоты.
Структура билирубина и уробилиногена похожи, но одна двойная связь заменяется на одинарную в молекуле уробилиногена.
Большая часть уробилиногена, около 90% превращается в стеркобилин.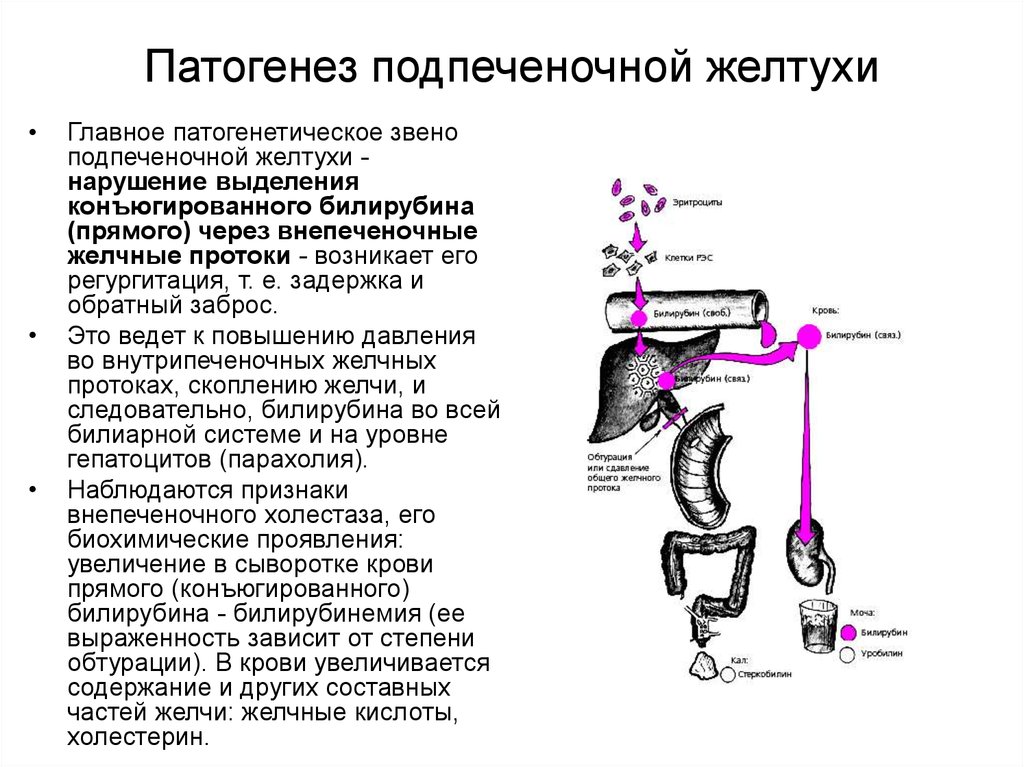
Который придаёт калу коричневый цвет.
Меньшая часть уробилиногена, около 10% подвергается реабсорбции в кишечнике.
Уробилиноген по системе воротной вены доставляется в печень. 5% превращается в конъюгированный билирубин. Часть уробилиногена превращается в уробилин и выделяется с мочой, придавая ей жёлтое окрашивание.
Неконъюгированный билирубин – липид растворимый Конъюгированный билирубин – воднорастворимый.
Желчь
Билирубин – выводится из организма.
Синтез и выделение билирубина, клиническая оценка:
В сыворотке крови определяется:
Уробилиноген мочи (в норме небольшое количество)
Билирубин в моче (конъюгированный – в норме отсутствует).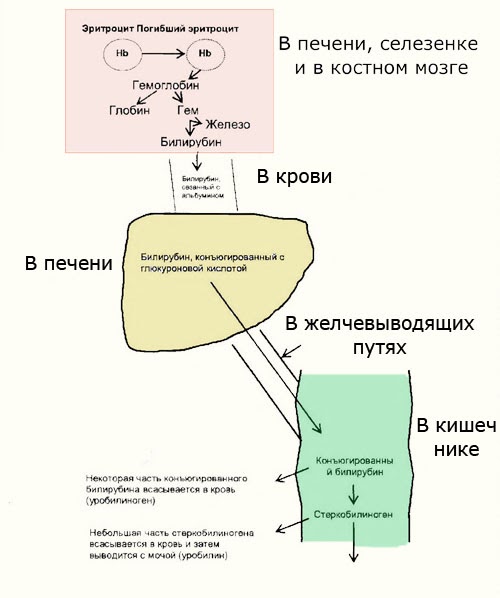
4 общие причины повышения билирубина:
При гемолизе повышается уровень непрямого билирубина, при обструкции прямого, при заболеваниях печени – повышаются уровни как конъюгированного, так и неконъюгированного билирубина.
При повышении уровня билирубина важно оценить – повышение билирубина изолированное, или имеется повышение уровня и других показателей работы печени- повышение печёночных ферментов, щелочной фосфатазы.
При изолированном повышении уровня билирубина исключаются специальные причины. При повышении печёночных ферментов, щелочной фосфатазы выделяют гепатоцеллюлярный и холестатический тип повреждения печени и проводится дальнейший дифференциальный диагноз для уточнения причины нарушения функции печени и проведения соответствующей терапии.
Симулятор клинического случая “Хронический миелолейкоз и желтуха”
Зарегистрируйтесь на нашем сайте прямо сейчас, чтобы иметь доступ к большему количеству обучающих материалов!
Подписаться на наши страницы:
Адитья Калаконда; Бьянка А. Дженкинс; Савио Джон.
Информация об авторе
Последнее обновление: 12 сентября 2022 г.
Билирубин является важным метаболитом гема (ферропротопорфирин IX), координационным комплексом, который служит для координации железа в различных белках. Это потенциально токсичное вещество. Однако в организме выработаны механизмы его безопасной детоксикации и утилизации.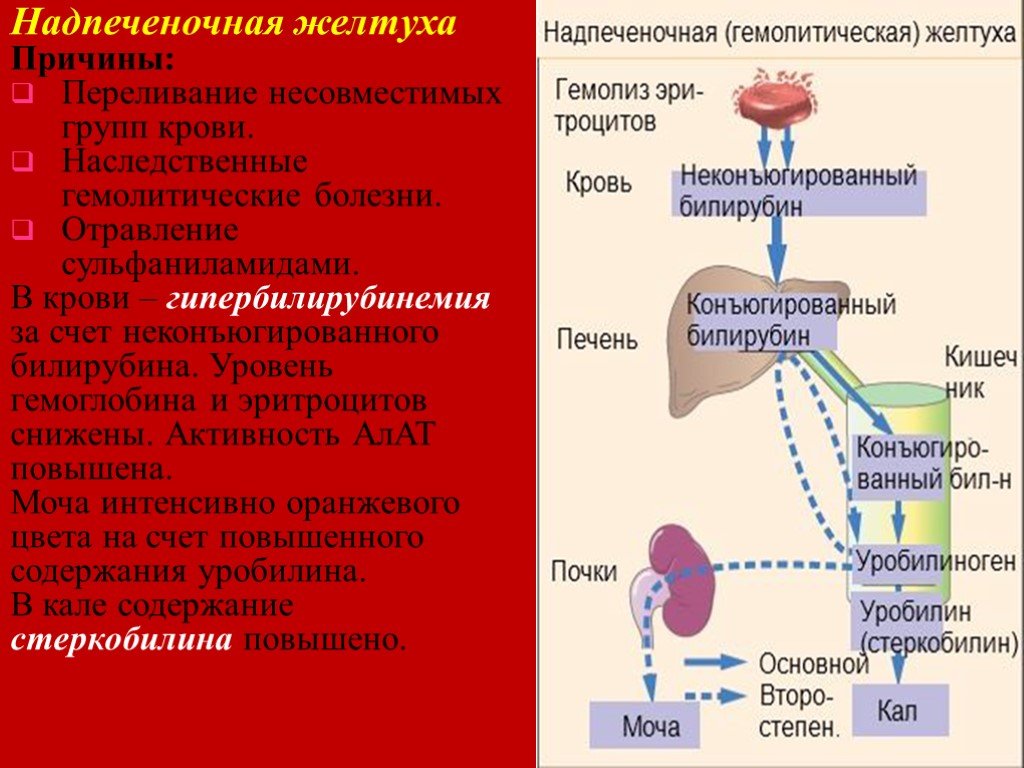 Билирубин и его метаболиты также придают характерный желтый цвет желчи и стулу и, в меньшей степени, моче. В этой статье будет обобщен механизм метаболизма гема и синтеза билирубина.[1][2][3]
Билирубин и его метаболиты также придают характерный желтый цвет желчи и стулу и, в меньшей степени, моче. В этой статье будет обобщен механизм метаболизма гема и синтеза билирубина.[1][2][3]
Образование билирубина
Билирубин получают из двух основных источников. Примерно 80% билирубина образуется в результате распада гемоглобина в стареющих эритроцитах и преждевременно разрушенных эритроидных клеток в костном мозге. Остальная часть образуется в результате оборота различных гемсодержащих белков, обнаруженных в других тканях, прежде всего в печени и мышцах. Эти белки включают миоглобин, цитохромы, каталазу, пероксидазу и триптофанпирролазу. Ежедневно вырабатывается около 4 мг/кг массы тела билирубина.
Метаболизм клеточного гема
Гем представляет собой кольцо из четырех пирролов, соединенных углеродными мостиками и центральным атомом железа. Билирубин образуется в результате двухстадийной последовательной каталитической реакции деградации, которая в основном происходит в клетках ретикулоэндотелиальной системы, особенно в селезенке.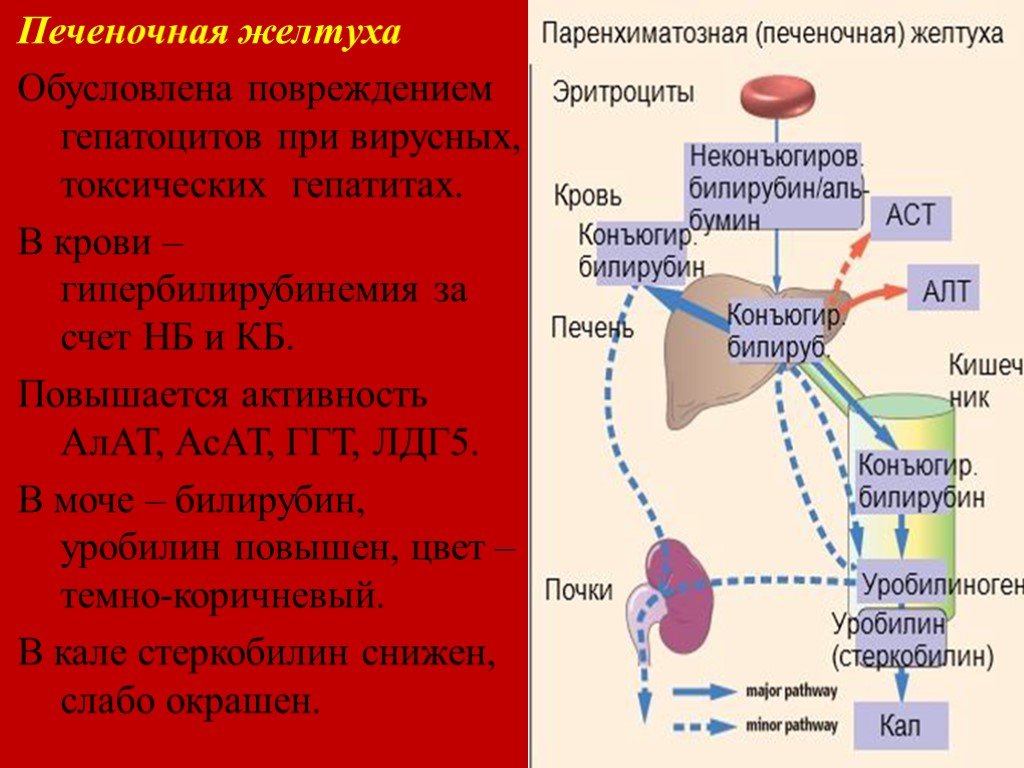 Другие клетки включают фагоциты и клетки Купфера печени. Гем поглощается Эти клетки поглощают гем, и на них действует фермент гемоксигеназа. Фермент высвобождает хелатное железо, катализируя окисление альфа-углеродного мостика. В результате этой реакции образуется эквимолярное количество окиси углерода, которая выделяется легкими и приводит к образованию зеленого пигмента биливердина. На этот зеленый пигмент дополнительно воздействует зависимый от никотинамидадениндинуклеотидфосфата (НАДФН) фермент, биливердинредуктаза. Этот процесс высвобождает оранжево-желтый пигмент, известный как билирубин. Гемоксигеназа, как упоминалось выше, присутствует в высоких концентрациях в клетках Купфера печени и клетках ретикулоэндотелиальной системы. Гемоксигеназа является фактором, ограничивающим скорость образования билирубина.
Другие клетки включают фагоциты и клетки Купфера печени. Гем поглощается Эти клетки поглощают гем, и на них действует фермент гемоксигеназа. Фермент высвобождает хелатное железо, катализируя окисление альфа-углеродного мостика. В результате этой реакции образуется эквимолярное количество окиси углерода, которая выделяется легкими и приводит к образованию зеленого пигмента биливердина. На этот зеленый пигмент дополнительно воздействует зависимый от никотинамидадениндинуклеотидфосфата (НАДФН) фермент, биливердинредуктаза. Этот процесс высвобождает оранжево-желтый пигмент, известный как билирубин. Гемоксигеназа, как упоминалось выше, присутствует в высоких концентрациях в клетках Купфера печени и клетках ретикулоэндотелиальной системы. Гемоксигеназа является фактором, ограничивающим скорость образования билирубина.
Окончательная структура сильно уплотнена за счет водородных связей, что делает молекулу практически нерастворимой в водных растворах при нейтральном pH. Полностью связанная структура билирубина обозначается как билирубин IX-альфа-ZZ.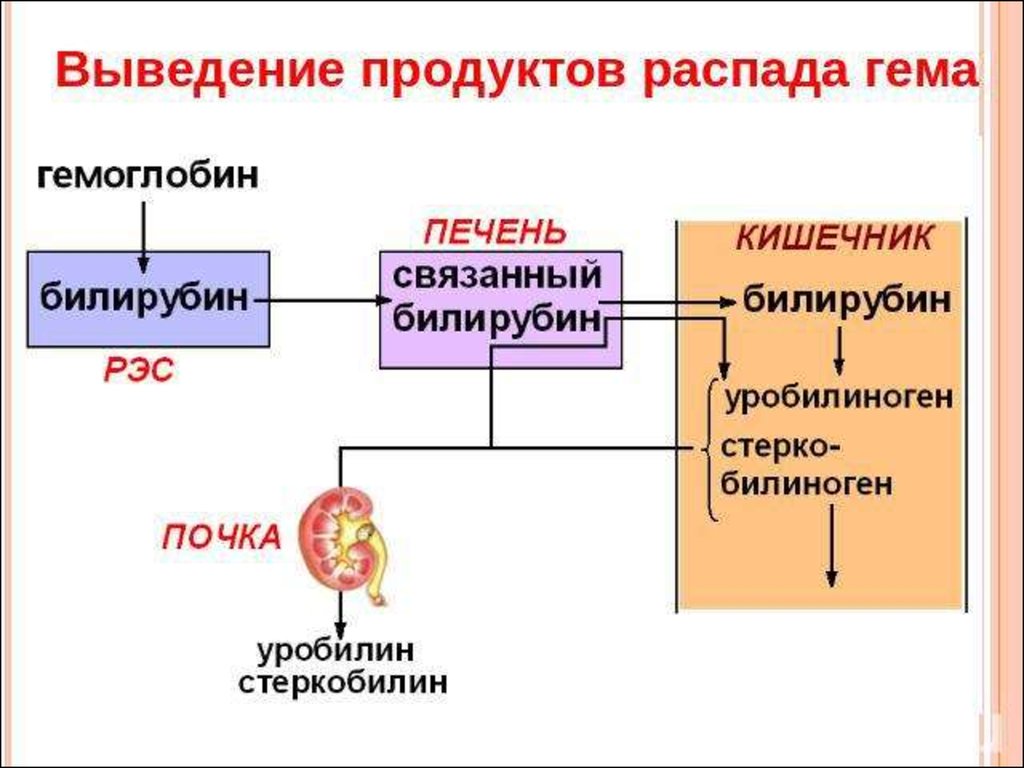 Билирубин, будучи нерастворимым в водном растворе, переносится в кровотоке в связи с альбумином, что представляет собой обратимую и ковалентную связь.
Билирубин, будучи нерастворимым в водном растворе, переносится в кровотоке в связи с альбумином, что представляет собой обратимую и ковалентную связь.
Метаболизм билирубина
Связывание с альбумином: Как только билирубин попадает в плазму, он поглощается альбумином, который служит его переносчиком по всему телу. Сродство связывания альбумина с билирубином чрезвычайно велико, и в идеальных условиях в плазме не наблюдается свободного (несвязанного с альбумином) неконъюгированного билирубина. В меньшей степени, особенно при состояниях гипоальбуминемии, связывание также происходит с липопротеинами высокой плотности. Связывание альбумина ограничивает выход билирубина из сосудистого пространства, сводит к минимуму клубочковую фильтрацию и препятствует его преципитации и отложению в тканях. Когда альбумин-билирубиновый комплекс достигает печени, высокопроницаемая печеночная циркуляция позволяет комплексу достичь синусоидальной поверхности гепатоцита.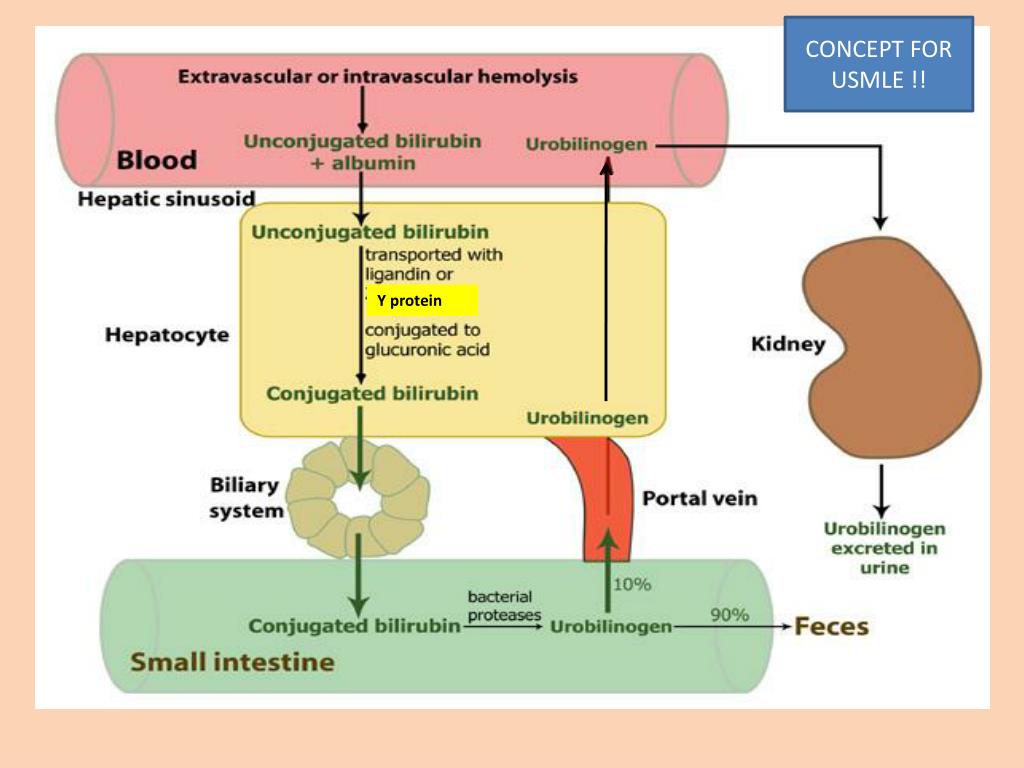 Это позволяет пигменту отделиться от альбумина и попасть в печень. Этот процесс относительно неэффективен, так как клиренс билирубина при первом прохождении составляет примерно 20%. Этот неэффективный процесс позволяет всегда иметь возможность измерить концентрацию неконъюгированного билирубина, связанного с альбумином, в венозном кровотоке. Связывание альбумина с билирубином обратимо.
Это позволяет пигменту отделиться от альбумина и попасть в печень. Этот процесс относительно неэффективен, так как клиренс билирубина при первом прохождении составляет примерно 20%. Этот неэффективный процесс позволяет всегда иметь возможность измерить концентрацию неконъюгированного билирубина, связанного с альбумином, в венозном кровотоке. Связывание альбумина с билирубином обратимо.
Механизмы печеночного транспорта: Билирубин поступает в гепатоциты из синусоидов печени по двум различным механизмам: пассивная диффузия и рецептор-опосредованный эндоцитоз. Процесс пассивной диффузии не требует больших затрат энергии и, как следствие, следует градиенту концентрации, что делает поток двунаправленным. Активный перенос неконъюгированного билирубина из печеночных синусоидов опосредуется белками-переносчиками, которые недостаточно изучены. Большая часть неконъюгированного билирубина, поступающего в гепатоциты, экстрагируется в перипортальной области. Часть конъюгированного и неконъюгированного билирубина в гепатоците транспортируется обратно в синусоидальное пространство, и эта фракция снова поднимается вниз по течению в синусоидальный поток.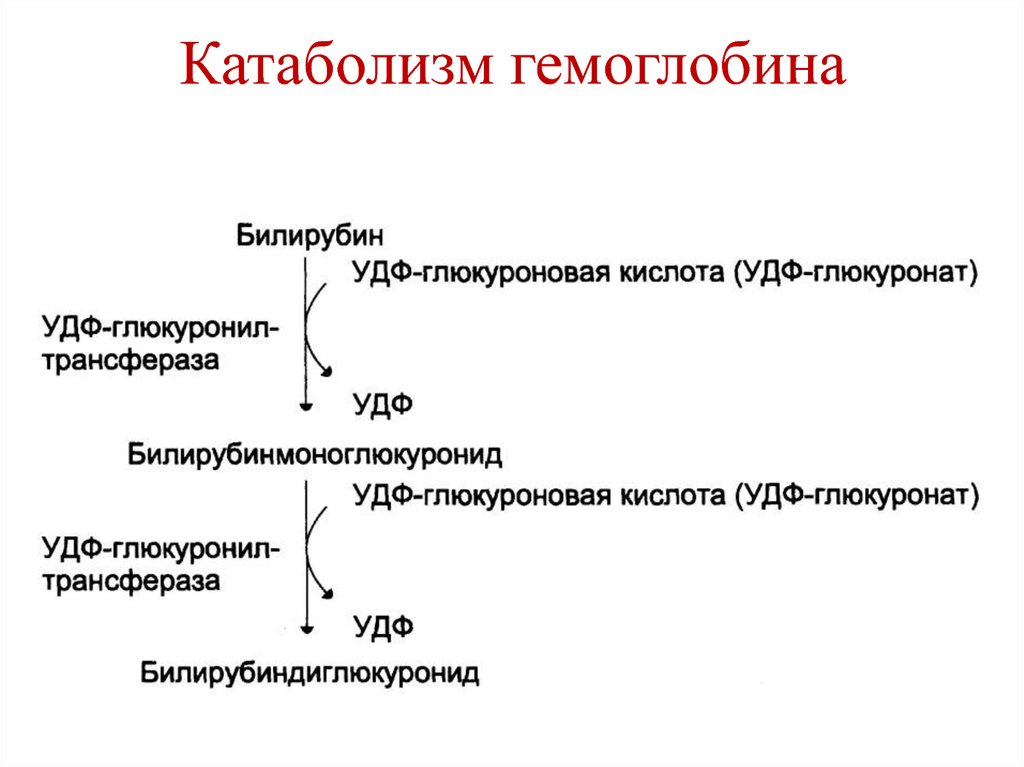 Поглощение опосредовано членами 1A и 1B семейства полипептидов, переносящих органические анионы (OATP). Эти полипептиды кодируются генами: SLCO1B1 и SLCO1B3 . Связанный билирубин, который избегает обратного захвата гепатоцитами, выводится с мочой. Связывание билирубина с глутатион-S-трансферазами, которое само по себе увеличивает чистое поглощение, минимизирует отток интернализованного билирубина.
Поглощение опосредовано членами 1A и 1B семейства полипептидов, переносящих органические анионы (OATP). Эти полипептиды кодируются генами: SLCO1B1 и SLCO1B3 . Связанный билирубин, который избегает обратного захвата гепатоцитами, выводится с мочой. Связывание билирубина с глутатион-S-трансферазами, которое само по себе увеличивает чистое поглощение, минимизирует отток интернализованного билирубина.
Конъюгация гепатоцитов
Конъюгация обязательна для превращения билирубина в растворимую в воде и облегчения его секреции через канальцевую мембрану и экскреции с желчью. Билирубин конъюгируется в гепатоцитах с глюкуроновой кислотой с помощью семейства ферментов, называемых уридин-дифосфоглюкуроновой глюкуронозилтрансферазой (УДФГТ). Процесс глюкуронирования является одним из многих важнейших механизмов детоксикации организма человека. Существует множество различных изоформ UDPGT, но физиологически важной изоформой для глюкуронирования билирубина является UDPGT1A1.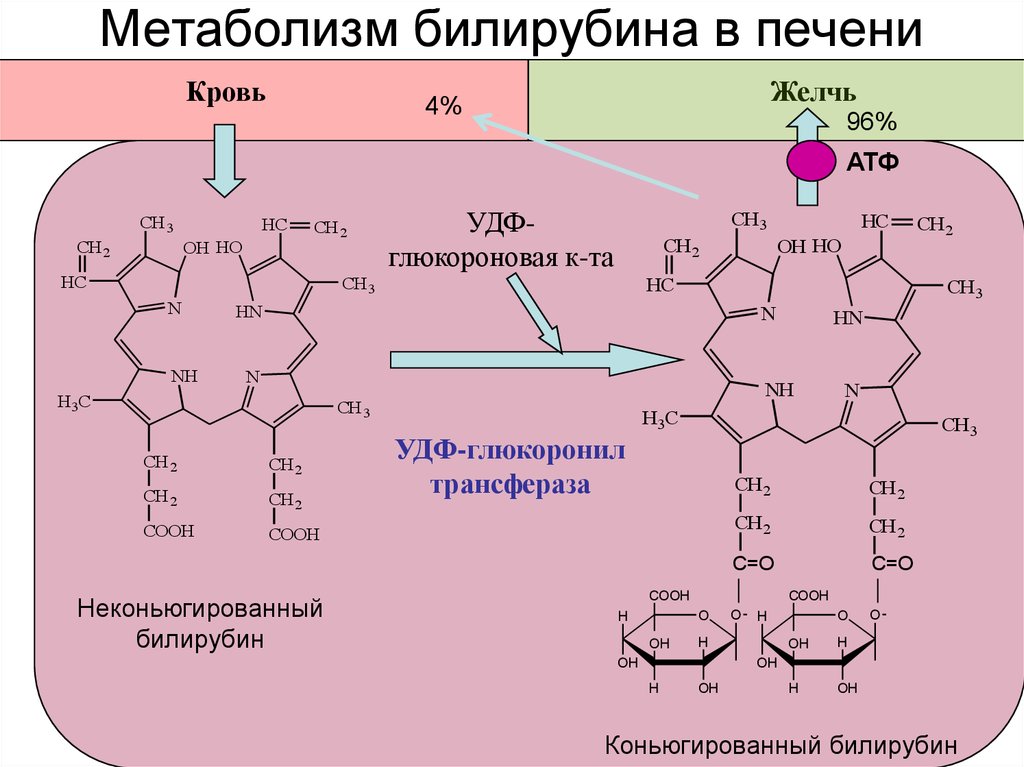 Фермент этерифицирует два фрагмента глюкуронида с боковыми цепями пропионовой кислоты билирубина. В нормальных условиях преобладающей синтезируемой молекулой является диглюкуронид билирубина. Однако, если система конъюгации перегружена в условиях избыточного синтеза билирубина, большая часть билирубина может быть конъюгирована в виде моноглюкуронида билирубина. Соотношение моноконъюгированного пигмента к дисконъюгированному в желчи составляет 1:4. Конъюгация билирубина в водорастворимую форму включает разрушение водородных связей, что является важным процессом для его выведения печенью и почками. Это достигается путем конъюгации глюкуроновой кислотой боковых цепей пропионовой кислоты билирубина.
Фермент этерифицирует два фрагмента глюкуронида с боковыми цепями пропионовой кислоты билирубина. В нормальных условиях преобладающей синтезируемой молекулой является диглюкуронид билирубина. Однако, если система конъюгации перегружена в условиях избыточного синтеза билирубина, большая часть билирубина может быть конъюгирована в виде моноглюкуронида билирубина. Соотношение моноконъюгированного пигмента к дисконъюгированному в желчи составляет 1:4. Конъюгация билирубина в водорастворимую форму включает разрушение водородных связей, что является важным процессом для его выведения печенью и почками. Это достигается путем конъюгации глюкуроновой кислотой боковых цепей пропионовой кислоты билирубина.
Экскреция конъюгированной желчи: Конъюгированный билирубин и другие вещества, предназначенные для экскреции с желчью, активно транспортируются через мембрану желчных канальцев гепатоцита. Градиент концентрации очень высок и может достигать 1:1000. Известно по крайней мере четыре канальцевых транспортера, которые участвуют в экскреции конъюгированного билирубина.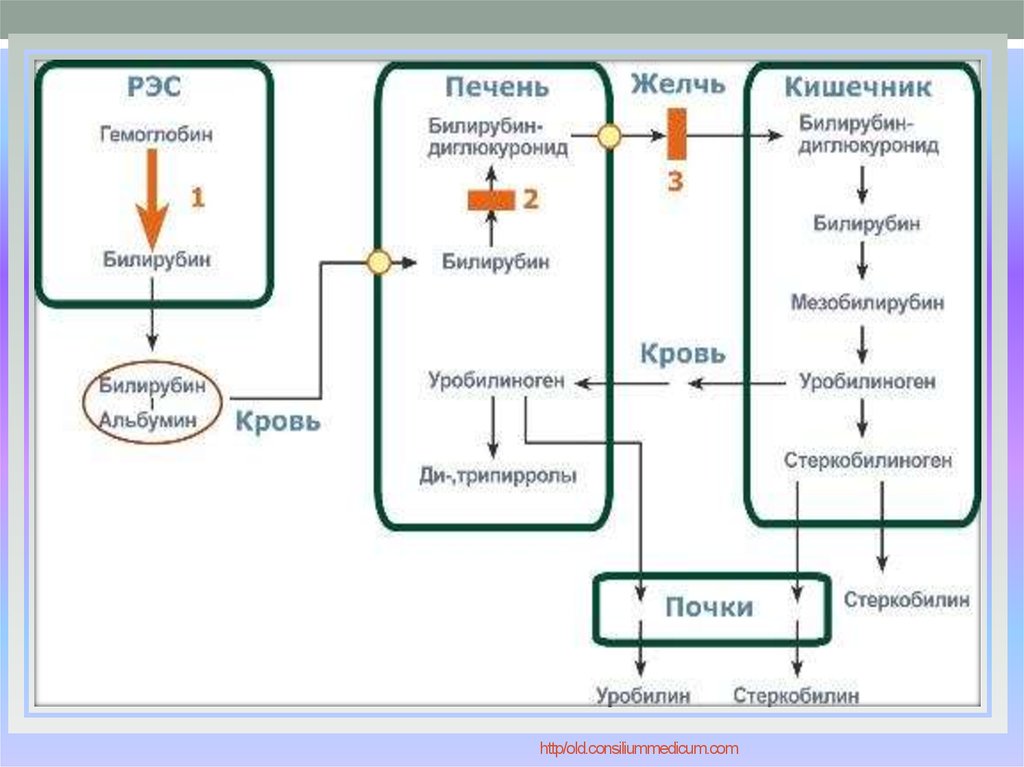 Тем не менее, белок 2, ассоциированный с множественной лекарственной устойчивостью (MRP2), по-видимому, играет доминирующую роль в канальцевой секреции конъюгированного билирубина. Часть конъюгированного билирубина транспортируется в синусоиды и портальную циркуляцию с помощью MRP3, который может подвергаться обратному захвату гепатоцитами через синусоидальные белки, белки транспорта органических анионов 1B1 и 1B3 (OATP1B1 и OATP1B3). Таким образом, некоторое количество конъюгированного и неконъюгированного билирубина может выходить из цитозоля гепатоцитов в плазму, где он связывается с альбумином и транспортируется по всему телу. Однако в желчь может поступать только конъюгированный билирубин. Затем конъюгированный билирубин активно секретируется в канальцевую желчь и стекает в тонкую кишку. Ступенью, ограничивающей скорость выделения билирубина, является экскреторная способность печени по конъюгированному билирубину. Часть конъюгированного билирубина может накапливаться в сыворотке крови при нарушении печеночной экскреции конъюгированного билирубина, например, при длительной билиарной обструкции или внутрипеченочном холестазе.
Тем не менее, белок 2, ассоциированный с множественной лекарственной устойчивостью (MRP2), по-видимому, играет доминирующую роль в канальцевой секреции конъюгированного билирубина. Часть конъюгированного билирубина транспортируется в синусоиды и портальную циркуляцию с помощью MRP3, который может подвергаться обратному захвату гепатоцитами через синусоидальные белки, белки транспорта органических анионов 1B1 и 1B3 (OATP1B1 и OATP1B3). Таким образом, некоторое количество конъюгированного и неконъюгированного билирубина может выходить из цитозоля гепатоцитов в плазму, где он связывается с альбумином и транспортируется по всему телу. Однако в желчь может поступать только конъюгированный билирубин. Затем конъюгированный билирубин активно секретируется в канальцевую желчь и стекает в тонкую кишку. Ступенью, ограничивающей скорость выделения билирубина, является экскреторная способность печени по конъюгированному билирубину. Часть конъюгированного билирубина может накапливаться в сыворотке крови при нарушении печеночной экскреции конъюгированного билирубина, например, при длительной билиарной обструкции или внутрипеченочном холестазе.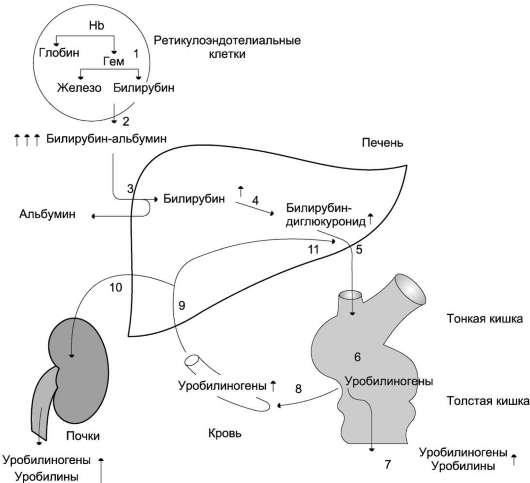 Эта фракция конъюгированного билирубина ковалентно связывается с альбумином и называется дельта-билирубином или дельта-фракцией или билипротеином. Поскольку дельта-билирубин связан с альбумином, его выведение из сыворотки занимает около 12-14 дней (что соответствует периоду полувыведения альбумина) в отличие от обычных 2-4 часов (период полувыведения билирубина).
Эта фракция конъюгированного билирубина ковалентно связывается с альбумином и называется дельта-билирубином или дельта-фракцией или билипротеином. Поскольку дельта-билирубин связан с альбумином, его выведение из сыворотки занимает около 12-14 дней (что соответствует периоду полувыведения альбумина) в отличие от обычных 2-4 часов (период полувыведения билирубина).
Процесс конъюгации изменяет физико-химические свойства билирубина, придавая ему множество особых свойств. Самое главное, он делает молекулу водорастворимой, что позволяет ей транспортироваться в желчи без белкового носителя. Конъюгация также увеличивает размер молекулы. Конъюгация предотвращает пассивную реабсорбцию билирубина слизистой оболочкой кишечника из-за его гидрофильности и большого молекулярного размера. Таким образом, конъюгация способствует элиминации потенциально токсичных продуктов метаболизма. Кроме того, конъюгация незначительно снижает сродство билирубина к альбумину.
Разложение в пищеварительном тракте: конъюгированный билирубин не реабсорбируется из проксимального отдела кишечника, как указано выше; для сравнения, неконъюгированный билирубин частично реабсорбируется через липидную мембрану эпителия тонкой кишки и подвергается энтерогепатической циркуляции. В проксимальном отделе тонкой кишки не происходит дополнительного метаболизма билирубина и происходит очень небольшая деконъюгация. Напротив, когда конъюгированный билирубин достигает дистального отдела подвздошной кишки и толстой кишки, он быстро восстанавливается и деконъюгируется кишечной флорой до ряда молекул, называемых уробилиногеном. Основные уробилиноиды, обнаруживаемые в стуле, известны как уробилиноген и стеркобилиноген, природа и относительная пропорция которых будут зависеть от наличия и состава кишечной бактериальной флоры. Эти вещества бесцветны, но после окисления до уробилина становятся оранжево-желтыми, что придает калу характерный цвет.
В проксимальном отделе тонкой кишки не происходит дополнительного метаболизма билирубина и происходит очень небольшая деконъюгация. Напротив, когда конъюгированный билирубин достигает дистального отдела подвздошной кишки и толстой кишки, он быстро восстанавливается и деконъюгируется кишечной флорой до ряда молекул, называемых уробилиногеном. Основные уробилиноиды, обнаруживаемые в стуле, известны как уробилиноген и стеркобилиноген, природа и относительная пропорция которых будут зависеть от наличия и состава кишечной бактериальной флоры. Эти вещества бесцветны, но после окисления до уробилина становятся оранжево-желтыми, что придает калу характерный цвет.
Измерение билирубина в сыворотке
Билирубин сыворотки измеряется спектрофотометрически, когда молекула подвергается реакции с диазореагентами, вызывающей расщепление тетрапиррола на два азодипиррола. Эта реакция получила название реакции Ван ден Берга. Неконъюгированный билирубин медленно реагирует с диазореагентом, поскольку центральный углеродный мостик билирубина скрыт внутри водородных связей.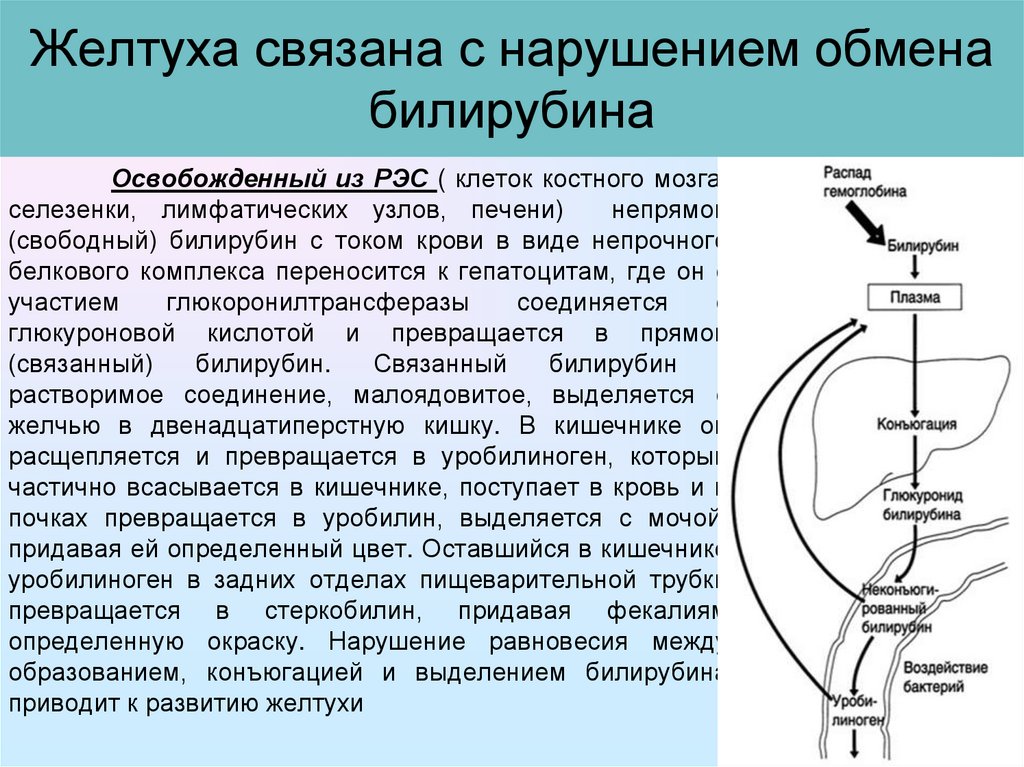 Напротив, в конъюгированном билирубине эти водородные связи отсутствуют, и реакция протекает быстро даже в отсутствие ускорителей. Добавление ускорителей, таких как кофеин или метанол, разрушает водородные связи, и реакция быстро завершается, давая значение общего билирубина. Неконъюгированный билирубин измеряется путем вычитания доли прямой реакции из общего билирубина. Потенциальные источники ошибок включают липиды плазмы, такие лекарства, как пропранолол и некоторые другие эндогенные вещества. Они мешают диазоанализу и потенциально могут дать ненадежный результат.
Напротив, в конъюгированном билирубине эти водородные связи отсутствуют, и реакция протекает быстро даже в отсутствие ускорителей. Добавление ускорителей, таких как кофеин или метанол, разрушает водородные связи, и реакция быстро завершается, давая значение общего билирубина. Неконъюгированный билирубин измеряется путем вычитания доли прямой реакции из общего билирубина. Потенциальные источники ошибок включают липиды плазмы, такие лекарства, как пропранолол и некоторые другие эндогенные вещества. Они мешают диазоанализу и потенциально могут дать ненадежный результат.
Поскольку неконъюгированный билирубин всегда связан с альбумином в сыворотке, он не может фильтроваться клубочками (при отсутствии гломерулярного заболевания). Таким образом, несвязанный билирубин никогда не обнаруживается в моче, даже если уровень несвязанного билирубина в крови повышен. Желтуха, возникающая при неконъюгированной гипербилирубинемии, называется ахолурической, поскольку моча не темнеет.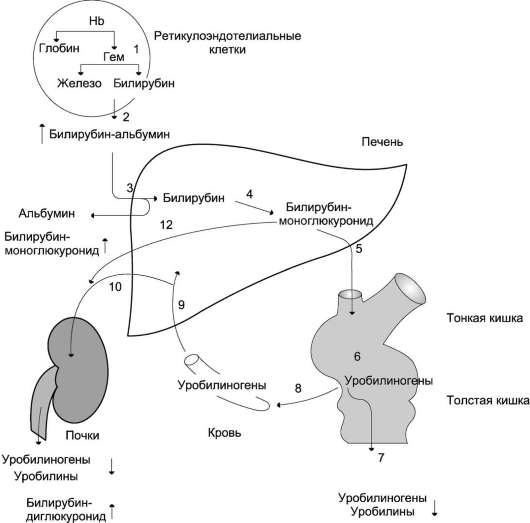 Однако темная моча возникает при экскреции избытка водорастворимого конъюгированного билирубина. Это наблюдается при конъюгированной гипербилирубинемии и указывает на наличие заболевания печени или желчевыводящих путей. Таким образом, присутствие билирубина в моче поможет идентифицировать тонкую гепатобилиарную дисфункцию, ведущую к конъюгированной гипербилирубинемии, даже если измеренная концентрация конъюгированного билирубина в сыворотке лишь незначительно повышена. Исключением из этого правила является отсутствие билирубинурии у больного с длительным холестазом и выраженной желтухой. Это связано с образованием дельта-билирубина или конъюгированного билирубина, который прочно связан с сывороточным альбумином. Отсутствие билирубинурии у таких пациентов не должно вызывать каких-либо трудностей при диагностике конъюгированной гипербилирубинемии, поскольку у пациента явно выражена желтуха, а конъюгированный билирубин в сыворотке в таких случаях заметно повышен.[6][7][8][9].][2]
Однако темная моча возникает при экскреции избытка водорастворимого конъюгированного билирубина. Это наблюдается при конъюгированной гипербилирубинемии и указывает на наличие заболевания печени или желчевыводящих путей. Таким образом, присутствие билирубина в моче поможет идентифицировать тонкую гепатобилиарную дисфункцию, ведущую к конъюгированной гипербилирубинемии, даже если измеренная концентрация конъюгированного билирубина в сыворотке лишь незначительно повышена. Исключением из этого правила является отсутствие билирубинурии у больного с длительным холестазом и выраженной желтухой. Это связано с образованием дельта-билирубина или конъюгированного билирубина, который прочно связан с сывороточным альбумином. Отсутствие билирубинурии у таких пациентов не должно вызывать каких-либо трудностей при диагностике конъюгированной гипербилирубинемии, поскольку у пациента явно выражена желтуха, а конъюгированный билирубин в сыворотке в таких случаях заметно повышен.[6][7][8][9].][2]
Получите бесплатный доступ к вопросам с несколькими вариантами ответов по этой теме.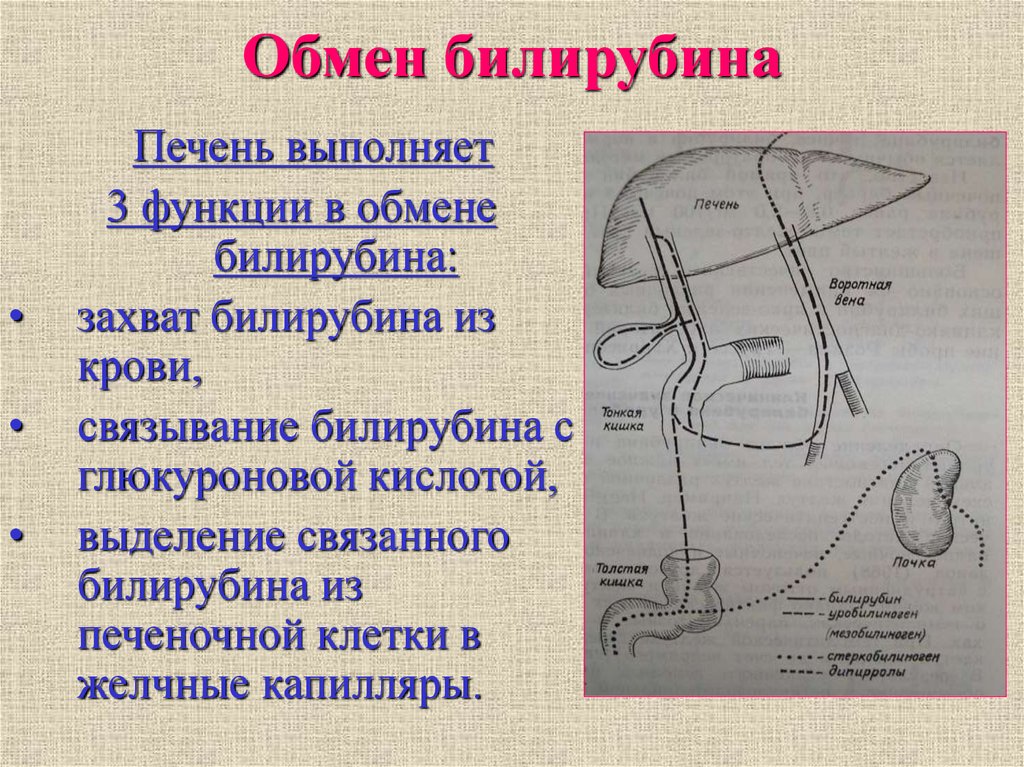
Комментарий к этой статье.
Метаболический путь билирубина в гепатоците. Билирубин-G соответствует глюкуронату билирубина, где донором является уридиндифосфат глюкуроновая кислота (УДФ-ГК). Это катализируется ферментом уридиндифосфат-глюкуронилтрансферазой (UGT1A1). Гилберт (подробнее...)
Космачевская О.В., Топунов А.Ф. Альтернативные и дополнительные функции гемоглобина эритроцитов. Биохимия (Москва). 2018 дек;83(12):1575-1593. [PubMed: 30878032]
Дош А.Р., Имагава Д.К., Ютрик З. Метаболизм желчи и литогенез: обновление. Surg Clin North Am. 2019 апр;99(2):215-229. [PubMed: 30846031]
Shen H, Zeng C, Wu X, Liu S, Chen X. Прогностическое значение общего билирубина у пациентов с острым инфарктом миокарда: метаанализ. Медицина (Балтимор). 2019Янв;98(3):e13920. [Бесплатная статья PMC: PMC6370131] [PubMed: 30653097]
Hinds TD, Stec DE. Билирубин, кардиометаболическая сигнальная молекула. Гипертония. 2018 окт; 72 (4): 788-795. [Бесплатная статья PMC: PMC6205727] [PubMed: 30354722]
Нгашангва Л., Бачу В., Госвами П. Разработка новых методов определения билирубина. Джей Фарм Биомед Анал. 2019 05 января; 162: 272-285. [PubMed: 30273817]
Бенешич А. [Лекарственное поражение печени (ЛПП)]. MMW Fortschr Med. 2019 май; 161(8):57-62. [PubMed: 31037662]
Patel SP, Vasavda C, Ho B, Meixiong J, Dong X, Kwatra SG. Холестатический зуд: новые механизмы и методы лечения. J Am Acad Дерматол. 2019 дек;81(6):1371-1378. [Бесплатная статья PMC: PMC7825249] [PubMed: 31009666]
Coucke EM, Akbar H, Kahloon A, Lopez PP. StatPearls [Интернет]. Издательство StatPearls; Остров сокровищ (Флорида): 26 сентября 2022 г. Непроходимость желчевыводящих путей. [В паблике: 30969520]

Снайдер Э., Кашьяп С., Лопес П.П. StatPearls [Интернет]. Издательство StatPearls; Остров сокровищ (Флорида): 25 июля 2022 г. Сканирование гепатобилиарной иминодиуксусной кислоты. [PubMed: 30969603]
Адитья Калаконда; Бьянка А. Дженкинс; Савио Джон.
Информация об авторе
Последнее обновление: 12 сентября 2022 г.
Билирубин является важным метаболитом гема (ферропротопорфирин IX), координационным комплексом, который служит для координации железа в различных белках. Это потенциально токсичное вещество. Однако в организме выработаны механизмы его безопасной детоксикации и утилизации. Билирубин и его метаболиты также придают характерный желтый цвет желчи и стулу и, в меньшей степени, моче. В этой статье будет обобщен механизм метаболизма гема и синтеза билирубина.[1][2][3]
Образование билирубина
Билирубин получают из двух основных источников.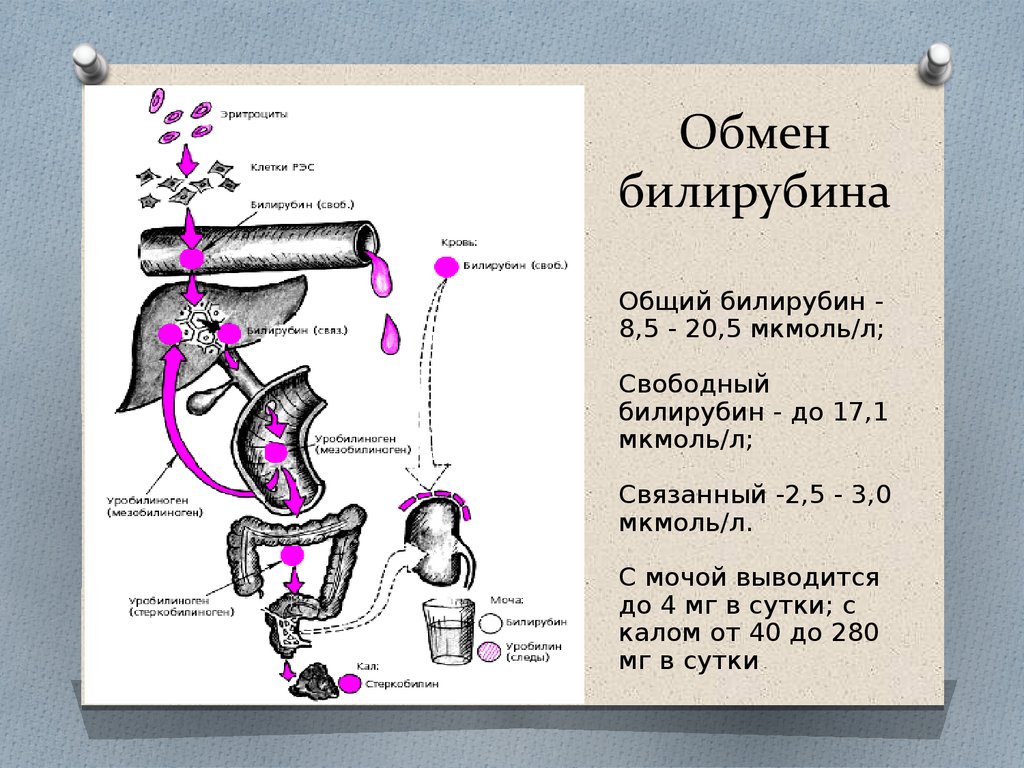 Примерно 80% билирубина образуется в результате распада гемоглобина в стареющих эритроцитах и преждевременно разрушенных эритроидных клеток в костном мозге. Остальная часть образуется в результате оборота различных гемсодержащих белков, обнаруженных в других тканях, прежде всего в печени и мышцах. Эти белки включают миоглобин, цитохромы, каталазу, пероксидазу и триптофанпирролазу. Ежедневно вырабатывается около 4 мг/кг массы тела билирубина.
Примерно 80% билирубина образуется в результате распада гемоглобина в стареющих эритроцитах и преждевременно разрушенных эритроидных клеток в костном мозге. Остальная часть образуется в результате оборота различных гемсодержащих белков, обнаруженных в других тканях, прежде всего в печени и мышцах. Эти белки включают миоглобин, цитохромы, каталазу, пероксидазу и триптофанпирролазу. Ежедневно вырабатывается около 4 мг/кг массы тела билирубина.
Метаболизм клеточного гема
Гем представляет собой кольцо из четырех пирролов, соединенных углеродными мостиками и центральным атомом железа. Билирубин образуется в результате двухстадийной последовательной каталитической реакции деградации, которая в основном происходит в клетках ретикулоэндотелиальной системы, особенно в селезенке. Другие клетки включают фагоциты и клетки Купфера печени. Гем поглощается Эти клетки поглощают гем, и на них действует фермент гемоксигеназа. Фермент высвобождает хелатное железо, катализируя окисление альфа-углеродного мостика.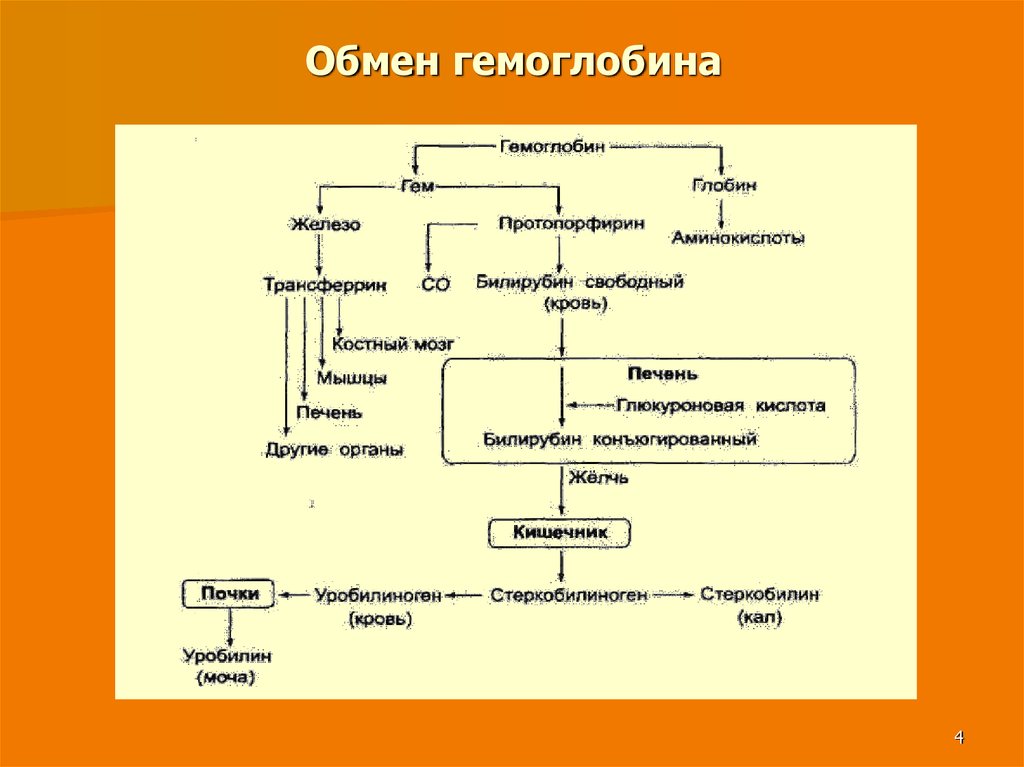 В результате этой реакции образуется эквимолярное количество окиси углерода, которая выделяется легкими и приводит к образованию зеленого пигмента биливердина. На этот зеленый пигмент дополнительно воздействует зависимый от никотинамидадениндинуклеотидфосфата (НАДФН) фермент, биливердинредуктаза. Этот процесс высвобождает оранжево-желтый пигмент, известный как билирубин. Гемоксигеназа, как упоминалось выше, присутствует в высоких концентрациях в клетках Купфера печени и клетках ретикулоэндотелиальной системы. Гемоксигеназа является фактором, ограничивающим скорость образования билирубина.
В результате этой реакции образуется эквимолярное количество окиси углерода, которая выделяется легкими и приводит к образованию зеленого пигмента биливердина. На этот зеленый пигмент дополнительно воздействует зависимый от никотинамидадениндинуклеотидфосфата (НАДФН) фермент, биливердинредуктаза. Этот процесс высвобождает оранжево-желтый пигмент, известный как билирубин. Гемоксигеназа, как упоминалось выше, присутствует в высоких концентрациях в клетках Купфера печени и клетках ретикулоэндотелиальной системы. Гемоксигеназа является фактором, ограничивающим скорость образования билирубина.
Окончательная структура сильно уплотнена за счет водородных связей, что делает молекулу практически нерастворимой в водных растворах при нейтральном pH. Полностью связанная структура билирубина обозначается как билирубин IX-альфа-ZZ. Билирубин, будучи нерастворимым в водном растворе, переносится в кровотоке в связи с альбумином, что представляет собой обратимую и ковалентную связь.
Метаболизм билирубина
Связывание с альбумином: Как только билирубин попадает в плазму, он поглощается альбумином, который служит его переносчиком по всему телу.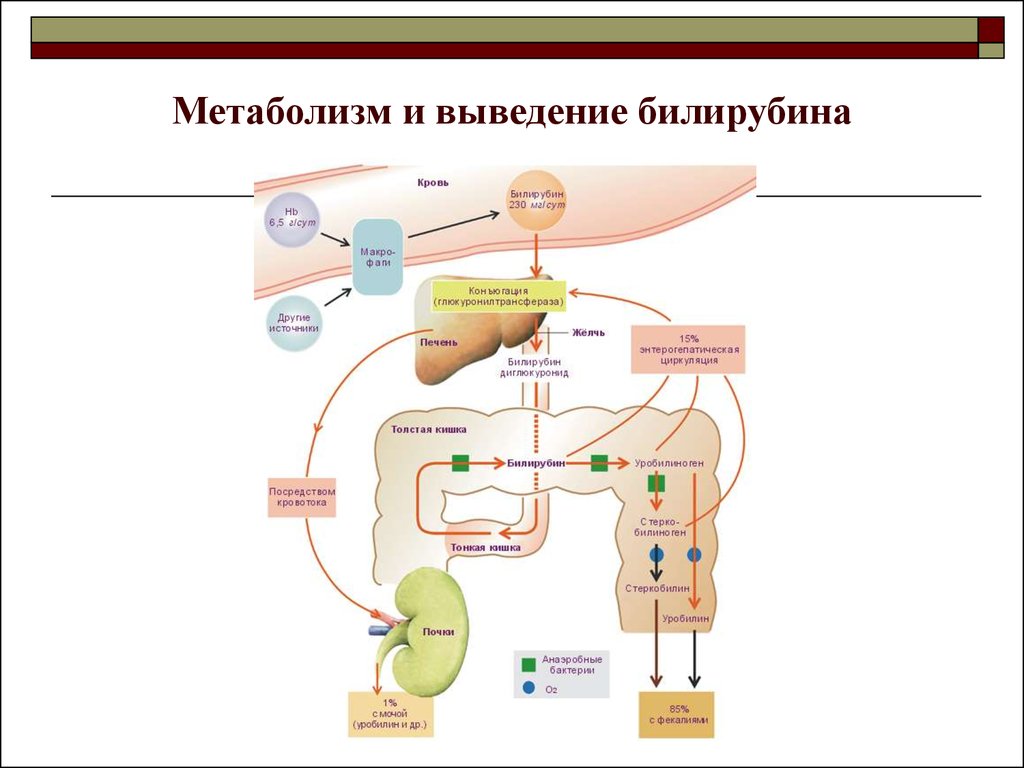 Сродство связывания альбумина с билирубином чрезвычайно велико, и в идеальных условиях в плазме не наблюдается свободного (несвязанного с альбумином) неконъюгированного билирубина. В меньшей степени, особенно при состояниях гипоальбуминемии, связывание также происходит с липопротеинами высокой плотности. Связывание альбумина ограничивает выход билирубина из сосудистого пространства, сводит к минимуму клубочковую фильтрацию и препятствует его преципитации и отложению в тканях. Когда альбумин-билирубиновый комплекс достигает печени, высокопроницаемая печеночная циркуляция позволяет комплексу достичь синусоидальной поверхности гепатоцита. Это позволяет пигменту отделиться от альбумина и попасть в печень. Этот процесс относительно неэффективен, так как клиренс билирубина при первом прохождении составляет примерно 20%. Этот неэффективный процесс позволяет всегда иметь возможность измерить концентрацию неконъюгированного билирубина, связанного с альбумином, в венозном кровотоке. Связывание альбумина с билирубином обратимо.
Сродство связывания альбумина с билирубином чрезвычайно велико, и в идеальных условиях в плазме не наблюдается свободного (несвязанного с альбумином) неконъюгированного билирубина. В меньшей степени, особенно при состояниях гипоальбуминемии, связывание также происходит с липопротеинами высокой плотности. Связывание альбумина ограничивает выход билирубина из сосудистого пространства, сводит к минимуму клубочковую фильтрацию и препятствует его преципитации и отложению в тканях. Когда альбумин-билирубиновый комплекс достигает печени, высокопроницаемая печеночная циркуляция позволяет комплексу достичь синусоидальной поверхности гепатоцита. Это позволяет пигменту отделиться от альбумина и попасть в печень. Этот процесс относительно неэффективен, так как клиренс билирубина при первом прохождении составляет примерно 20%. Этот неэффективный процесс позволяет всегда иметь возможность измерить концентрацию неконъюгированного билирубина, связанного с альбумином, в венозном кровотоке. Связывание альбумина с билирубином обратимо.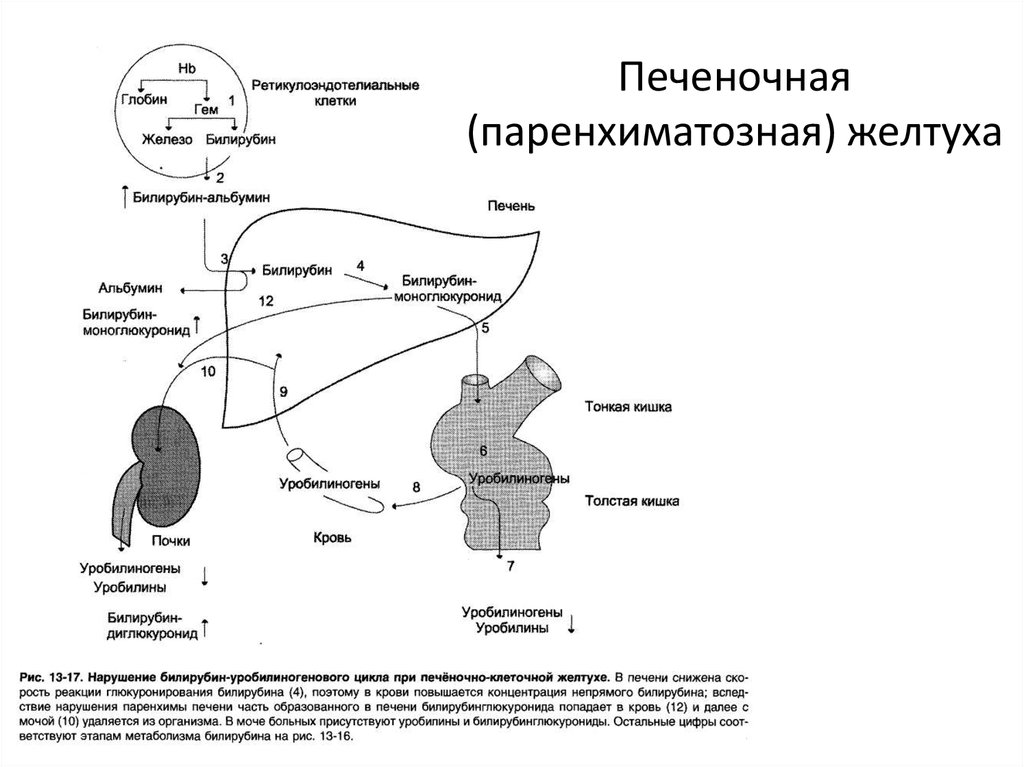
Механизмы печеночного транспорта: Билирубин поступает в гепатоциты из синусоидов печени по двум различным механизмам: пассивная диффузия и рецептор-опосредованный эндоцитоз. Процесс пассивной диффузии не требует больших затрат энергии и, как следствие, следует градиенту концентрации, что делает поток двунаправленным. Активный перенос неконъюгированного билирубина из печеночных синусоидов опосредуется белками-переносчиками, которые недостаточно изучены. Большая часть неконъюгированного билирубина, поступающего в гепатоциты, экстрагируется в перипортальной области. Часть конъюгированного и неконъюгированного билирубина в гепатоците транспортируется обратно в синусоидальное пространство, и эта фракция снова поднимается вниз по течению в синусоидальный поток. Поглощение опосредовано членами 1A и 1B семейства полипептидов, переносящих органические анионы (OATP). Эти полипептиды кодируются генами: SLCO1B1 и SLCO1B3 . Связанный билирубин, который избегает обратного захвата гепатоцитами, выводится с мочой.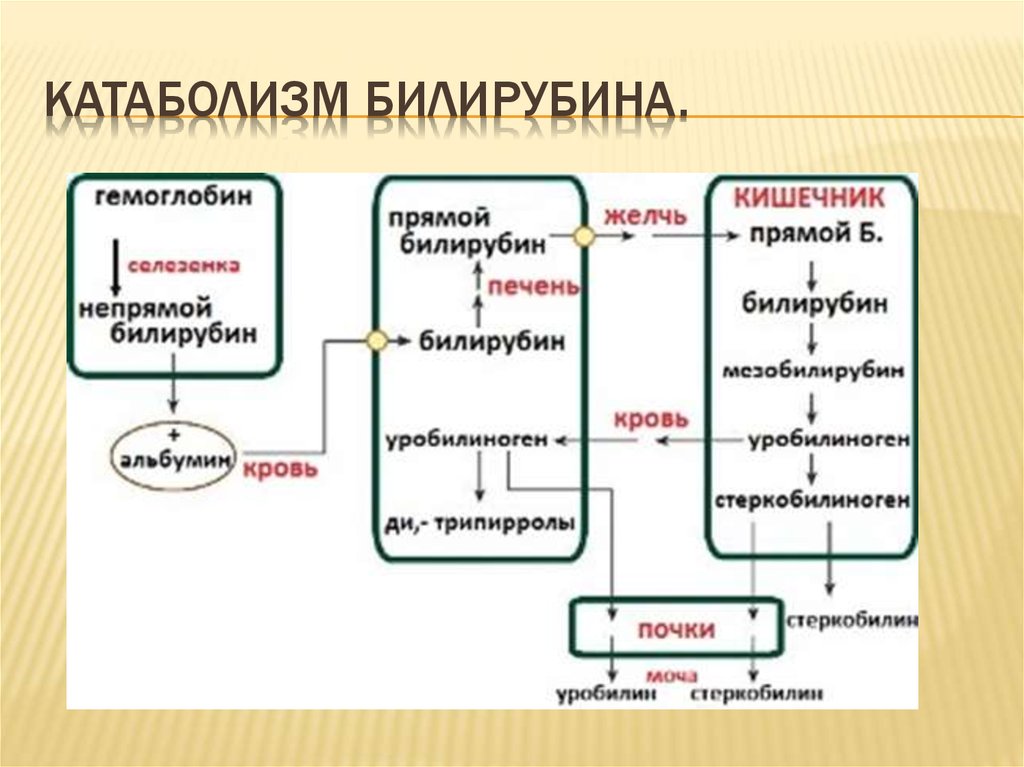 Связывание билирубина с глутатион-S-трансферазами, которое само по себе увеличивает чистое поглощение, минимизирует отток интернализованного билирубина.
Связывание билирубина с глутатион-S-трансферазами, которое само по себе увеличивает чистое поглощение, минимизирует отток интернализованного билирубина.
Конъюгация гепатоцитов
Конъюгация обязательна для превращения билирубина в растворимую в воде и облегчения его секреции через канальцевую мембрану и экскреции с желчью. Билирубин конъюгируется в гепатоцитах с глюкуроновой кислотой с помощью семейства ферментов, называемых уридин-дифосфоглюкуроновой глюкуронозилтрансферазой (УДФГТ). Процесс глюкуронирования является одним из многих важнейших механизмов детоксикации организма человека. Существует множество различных изоформ UDPGT, но физиологически важной изоформой для глюкуронирования билирубина является UDPGT1A1. Фермент этерифицирует два фрагмента глюкуронида с боковыми цепями пропионовой кислоты билирубина. В нормальных условиях преобладающей синтезируемой молекулой является диглюкуронид билирубина. Однако, если система конъюгации перегружена в условиях избыточного синтеза билирубина, большая часть билирубина может быть конъюгирована в виде моноглюкуронида билирубина. Соотношение моноконъюгированного пигмента к дисконъюгированному в желчи составляет 1:4. Конъюгация билирубина в водорастворимую форму включает разрушение водородных связей, что является важным процессом для его выведения печенью и почками. Это достигается путем конъюгации глюкуроновой кислотой боковых цепей пропионовой кислоты билирубина.
Соотношение моноконъюгированного пигмента к дисконъюгированному в желчи составляет 1:4. Конъюгация билирубина в водорастворимую форму включает разрушение водородных связей, что является важным процессом для его выведения печенью и почками. Это достигается путем конъюгации глюкуроновой кислотой боковых цепей пропионовой кислоты билирубина.
Экскреция конъюгированной желчи: Конъюгированный билирубин и другие вещества, предназначенные для экскреции с желчью, активно транспортируются через мембрану желчных канальцев гепатоцита. Градиент концентрации очень высок и может достигать 1:1000. Известно по крайней мере четыре канальцевых транспортера, которые участвуют в экскреции конъюгированного билирубина. Тем не менее, белок 2, ассоциированный с множественной лекарственной устойчивостью (MRP2), по-видимому, играет доминирующую роль в канальцевой секреции конъюгированного билирубина. Часть конъюгированного билирубина транспортируется в синусоиды и портальную циркуляцию с помощью MRP3, который может подвергаться обратному захвату гепатоцитами через синусоидальные белки, белки транспорта органических анионов 1B1 и 1B3 (OATP1B1 и OATP1B3). Таким образом, некоторое количество конъюгированного и неконъюгированного билирубина может выходить из цитозоля гепатоцитов в плазму, где он связывается с альбумином и транспортируется по всему телу. Однако в желчь может поступать только конъюгированный билирубин. Затем конъюгированный билирубин активно секретируется в канальцевую желчь и стекает в тонкую кишку. Ступенью, ограничивающей скорость выделения билирубина, является экскреторная способность печени по конъюгированному билирубину. Часть конъюгированного билирубина может накапливаться в сыворотке крови при нарушении печеночной экскреции конъюгированного билирубина, например, при длительной билиарной обструкции или внутрипеченочном холестазе. Эта фракция конъюгированного билирубина ковалентно связывается с альбумином и называется дельта-билирубином или дельта-фракцией или билипротеином. Поскольку дельта-билирубин связан с альбумином, его выведение из сыворотки занимает около 12-14 дней (что соответствует периоду полувыведения альбумина) в отличие от обычных 2-4 часов (период полувыведения билирубина).
Таким образом, некоторое количество конъюгированного и неконъюгированного билирубина может выходить из цитозоля гепатоцитов в плазму, где он связывается с альбумином и транспортируется по всему телу. Однако в желчь может поступать только конъюгированный билирубин. Затем конъюгированный билирубин активно секретируется в канальцевую желчь и стекает в тонкую кишку. Ступенью, ограничивающей скорость выделения билирубина, является экскреторная способность печени по конъюгированному билирубину. Часть конъюгированного билирубина может накапливаться в сыворотке крови при нарушении печеночной экскреции конъюгированного билирубина, например, при длительной билиарной обструкции или внутрипеченочном холестазе. Эта фракция конъюгированного билирубина ковалентно связывается с альбумином и называется дельта-билирубином или дельта-фракцией или билипротеином. Поскольку дельта-билирубин связан с альбумином, его выведение из сыворотки занимает около 12-14 дней (что соответствует периоду полувыведения альбумина) в отличие от обычных 2-4 часов (период полувыведения билирубина).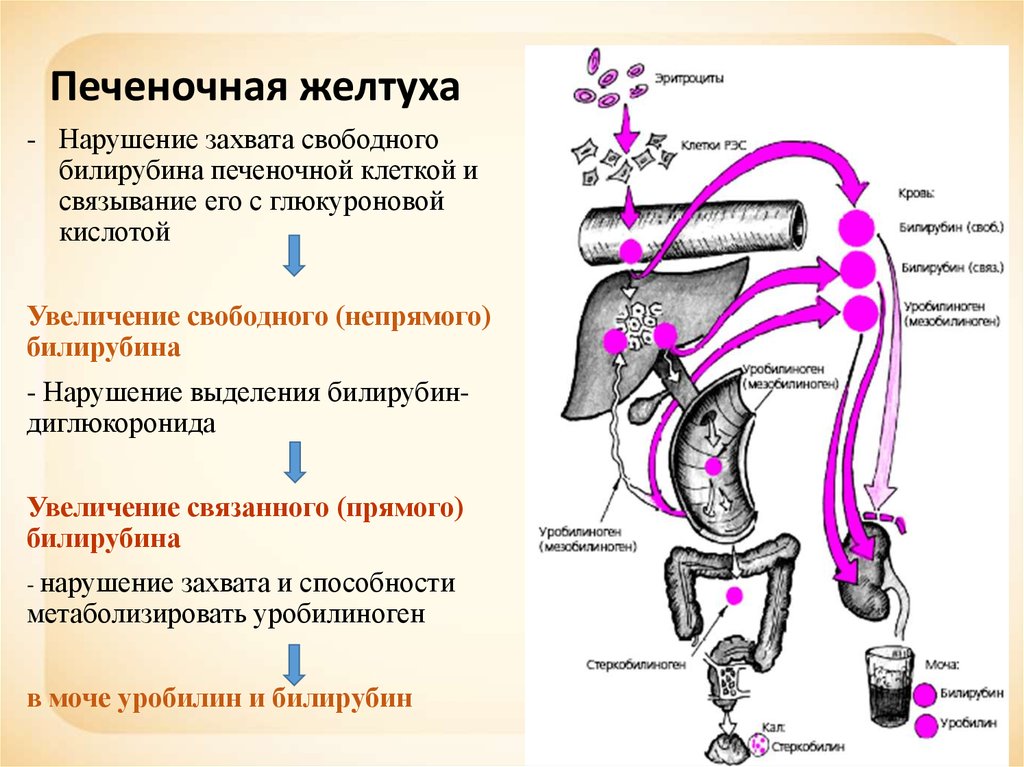
Процесс конъюгации изменяет физико-химические свойства билирубина, придавая ему множество особых свойств. Самое главное, он делает молекулу водорастворимой, что позволяет ей транспортироваться в желчи без белкового носителя. Конъюгация также увеличивает размер молекулы. Конъюгация предотвращает пассивную реабсорбцию билирубина слизистой оболочкой кишечника из-за его гидрофильности и большого молекулярного размера. Таким образом, конъюгация способствует элиминации потенциально токсичных продуктов метаболизма. Кроме того, конъюгация незначительно снижает сродство билирубина к альбумину.
Разложение в пищеварительном тракте: конъюгированный билирубин не реабсорбируется из проксимального отдела кишечника, как указано выше; для сравнения, неконъюгированный билирубин частично реабсорбируется через липидную мембрану эпителия тонкой кишки и подвергается энтерогепатической циркуляции. В проксимальном отделе тонкой кишки не происходит дополнительного метаболизма билирубина и происходит очень небольшая деконъюгация. Напротив, когда конъюгированный билирубин достигает дистального отдела подвздошной кишки и толстой кишки, он быстро восстанавливается и деконъюгируется кишечной флорой до ряда молекул, называемых уробилиногеном. Основные уробилиноиды, обнаруживаемые в стуле, известны как уробилиноген и стеркобилиноген, природа и относительная пропорция которых будут зависеть от наличия и состава кишечной бактериальной флоры. Эти вещества бесцветны, но после окисления до уробилина становятся оранжево-желтыми, что придает калу характерный цвет.
Напротив, когда конъюгированный билирубин достигает дистального отдела подвздошной кишки и толстой кишки, он быстро восстанавливается и деконъюгируется кишечной флорой до ряда молекул, называемых уробилиногеном. Основные уробилиноиды, обнаруживаемые в стуле, известны как уробилиноген и стеркобилиноген, природа и относительная пропорция которых будут зависеть от наличия и состава кишечной бактериальной флоры. Эти вещества бесцветны, но после окисления до уробилина становятся оранжево-желтыми, что придает калу характерный цвет.
Измерение билирубина в сыворотке
Билирубин сыворотки измеряется спектрофотометрически, когда молекула подвергается реакции с диазореагентами, вызывающей расщепление тетрапиррола на два азодипиррола. Эта реакция получила название реакции Ван ден Берга. Неконъюгированный билирубин медленно реагирует с диазореагентом, поскольку центральный углеродный мостик билирубина скрыт внутри водородных связей. Напротив, в конъюгированном билирубине эти водородные связи отсутствуют, и реакция протекает быстро даже в отсутствие ускорителей. Добавление ускорителей, таких как кофеин или метанол, разрушает водородные связи, и реакция быстро завершается, давая значение общего билирубина. Неконъюгированный билирубин измеряется путем вычитания доли прямой реакции из общего билирубина. Потенциальные источники ошибок включают липиды плазмы, такие лекарства, как пропранолол и некоторые другие эндогенные вещества. Они мешают диазоанализу и потенциально могут дать ненадежный результат.
Напротив, в конъюгированном билирубине эти водородные связи отсутствуют, и реакция протекает быстро даже в отсутствие ускорителей. Добавление ускорителей, таких как кофеин или метанол, разрушает водородные связи, и реакция быстро завершается, давая значение общего билирубина. Неконъюгированный билирубин измеряется путем вычитания доли прямой реакции из общего билирубина. Потенциальные источники ошибок включают липиды плазмы, такие лекарства, как пропранолол и некоторые другие эндогенные вещества. Они мешают диазоанализу и потенциально могут дать ненадежный результат.
Поскольку неконъюгированный билирубин всегда связан с альбумином в сыворотке, он не может фильтроваться клубочками (при отсутствии гломерулярного заболевания). Таким образом, несвязанный билирубин никогда не обнаруживается в моче, даже если уровень несвязанного билирубина в крови повышен. Желтуха, возникающая при неконъюгированной гипербилирубинемии, называется ахолурической, поскольку моча не темнеет. Однако темная моча возникает при экскреции избытка водорастворимого конъюгированного билирубина. Это наблюдается при конъюгированной гипербилирубинемии и указывает на наличие заболевания печени или желчевыводящих путей. Таким образом, присутствие билирубина в моче поможет идентифицировать тонкую гепатобилиарную дисфункцию, ведущую к конъюгированной гипербилирубинемии, даже если измеренная концентрация конъюгированного билирубина в сыворотке лишь незначительно повышена. Исключением из этого правила является отсутствие билирубинурии у больного с длительным холестазом и выраженной желтухой. Это связано с образованием дельта-билирубина или конъюгированного билирубина, который прочно связан с сывороточным альбумином. Отсутствие билирубинурии у таких пациентов не должно вызывать каких-либо трудностей при диагностике конъюгированной гипербилирубинемии, поскольку у пациента явно выражена желтуха, а конъюгированный билирубин в сыворотке в таких случаях заметно повышен.[6][7][8][9].][2]
Однако темная моча возникает при экскреции избытка водорастворимого конъюгированного билирубина. Это наблюдается при конъюгированной гипербилирубинемии и указывает на наличие заболевания печени или желчевыводящих путей. Таким образом, присутствие билирубина в моче поможет идентифицировать тонкую гепатобилиарную дисфункцию, ведущую к конъюгированной гипербилирубинемии, даже если измеренная концентрация конъюгированного билирубина в сыворотке лишь незначительно повышена. Исключением из этого правила является отсутствие билирубинурии у больного с длительным холестазом и выраженной желтухой. Это связано с образованием дельта-билирубина или конъюгированного билирубина, который прочно связан с сывороточным альбумином. Отсутствие билирубинурии у таких пациентов не должно вызывать каких-либо трудностей при диагностике конъюгированной гипербилирубинемии, поскольку у пациента явно выражена желтуха, а конъюгированный билирубин в сыворотке в таких случаях заметно повышен.[6][7][8][9].][2]
Получите бесплатный доступ к вопросам с несколькими вариантами ответов по этой теме.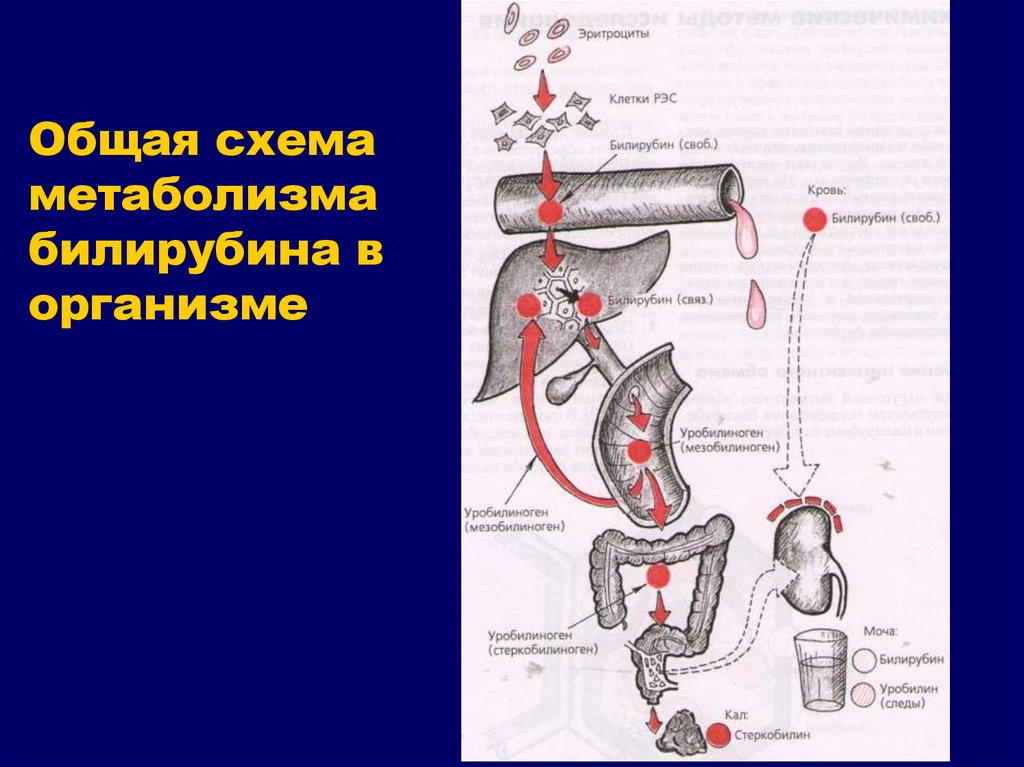
Комментарий к этой статье.
Метаболический путь билирубина в гепатоците. Билирубин-G соответствует глюкуронату билирубина, где донором является уридиндифосфат глюкуроновая кислота (УДФ-ГК). Это катализируется ферментом уридиндифосфат-глюкуронилтрансферазой (UGT1A1). Гилберт (подробнее...)
Космачевская О.В., Топунов А.Ф. Альтернативные и дополнительные функции гемоглобина эритроцитов. Биохимия (Москва). 2018 дек;83(12):1575-1593. [PubMed: 30878032]
Дош А.Р., Имагава Д.К., Ютрик З. Метаболизм желчи и литогенез: обновление. Surg Clin North Am. 2019 апр;99(2):215-229. [PubMed: 30846031]
Shen H, Zeng C, Wu X, Liu S, Chen X. Прогностическое значение общего билирубина у пациентов с острым инфарктом миокарда: метаанализ. Медицина (Балтимор). 2019Янв;98(3):e13920. [Бесплатная статья PMC: PMC6370131] [PubMed: 30653097]